Hay potenciales eléctricos a través de las membranas de prácticamente todas las células del cuerpo. Algunas células, como las células nerviosas y musculares, generan impulsos electroquímicos rápidamente cambiantes en sus membranas, y estos impulsos se utilizan para transmitir señales a través de las membranas de los nervios y de los músculos. En otros tipos de células, como las células glandulares, los macrófagos y las células ciliadas, los cambios locales de los potenciales de membrana también activan muchas de las funciones de las células.
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90 mV. Es decir, el potencial en el interior de la fibra es 90 mV más negativo que el potencial del líquido extracelular que está en el exterior de la misma, todas las membranas celulares del cuerpo tienen una potente bomba Na+-K+ que transporta continuamente iones sodio hacia el exterior de la célula e iones potasio hacia el interior se trata de una bomba electrógena porque se bombean más cargas positivas hacia el exterior que hacia el interior (tres iones Na+ hacia el exterior por cada dos iones K+ hacia el interior), dejando un déficit neto de iones positivos en el interior; esto genera un potencial negativo en el interior de la membrana celular.
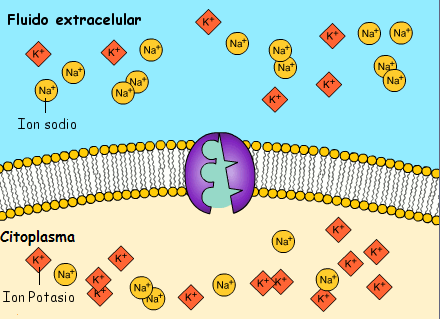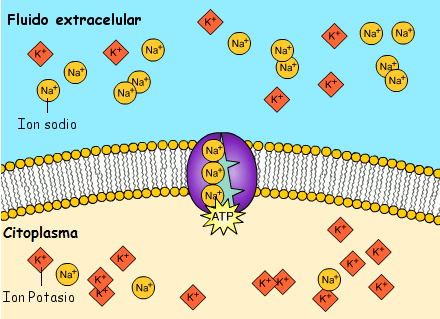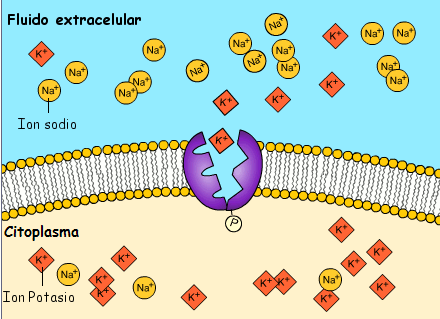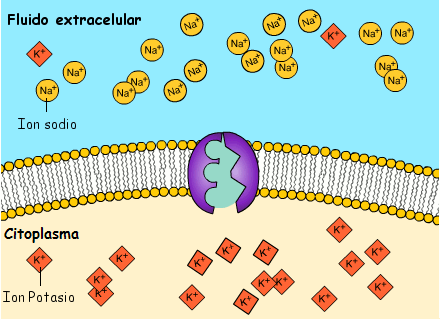
A veces el canal de potasio de dominios de poro en tándem, o canal de «fuga» de potasio (K+), en la membrana nerviosa a través dela que pueden escapar iones potasio incluso en una célula en reposo, stos canales de fuga de K+ también pueden dejar que se pierdan algunos iones sodio pero los canales son mucho más permeables al potasio que al sodio, en general aproximadamente 100 veces más permeables.
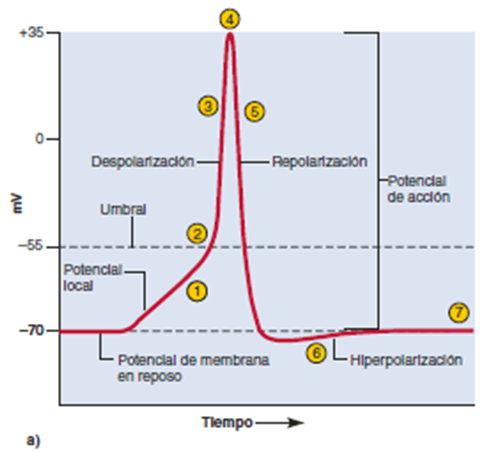
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega a su extremo. Las fases del potencial de acción son las siguientes:
Fase de reposo
La fase de reposo es el potencial de membrana en reposo antes del comienzo del potencial de acción. Se dice que la membrana está «polarizada» durante esta fase debido al potencial de membrana negativo de –90 mV que está presente.
Fase de despolarización
En este momento la membrana se hace súbitamente muy permeable a los iones sodio, lo que permite que un gran número de iones sodio con carga positiva difunda hacia el interior del axón. El estado «polarizado» normal de –90 mV se neutraliza inmediatamente por la entrada de iones sodio cargados positivamente, y el potencial aumenta rápidamente en dirección positiva, un proceso denominado despolarización. En las fibras nerviosas grandes el gran exceso de iones sodio positivos que se mueven hacia el interior hace que el potencial de membrana realmente se «sobreexcite» más allá del nivel cero y que se haga algo positivo. En algunas fibras más pequeñas, así como en muchas neuronas del sistema nervioso central, el potencial simplemente se acerca al nivel cero y no hay sobreexcitación hacia el estado positivo.
Fase de repolarización
En un plazo de algunas diezmilésimas de segundo después de que la membrana se haya hecho muy permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los canales de potasio se abren más de lo normal. De esta manera, la rápida difusión de los iones potasio hacia el exterior restablece el potencial de membrana en reposo negativo normal, que se denomina repolarización de la membrana.
El actor necesario en la producción tanto de la despolarización como de la repolarización de lamembrana nerviosa durante el potencial de acción es el canal de sodio activado por el voltaje. Uncanal de potasio activado por el voltaje también tiene una función importante en el aumento de la rapidez de la repolarización de la membrana. Estos dos canales activados por el voltaje tienen una función adicional a la de la bomba Na+-K+ y de los canales de fuga de K+.Este canal tiene dos compuertas, una cerca del exterior del canal, denominada compuerta de activación, y otra cerca del interior, denominada compuerta de inactivación. La parte superiorizquierda de la figura representa el estado de estas dos compuertas en la membrana en reposo normal,cuando el potencial de membrana es de –90 mV. En este estado la compuerta de activación estácerrada, lo que impide la entrada de iones sodio hacia el interior de la fibra a través de estos canalesde sodio.
Activación del canal de sodio
Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje (habitualmente algún punto entre –70 y –50 mV) que produce un cambio conformacional súbito en la activación de la compuerta, que bascula totalmente hasta la posición de abierta.
Inactivación del canal de sodio
El cambio conformacional que hace bascular la compuerta de inactivación hacia el estado cerrado es un proceso algo más lento que el cambio conformacional que abre la compuerta de activación. Por tanto, después de que el canal de sodio haya permanecido abierto durante algunas diezmilésimas de segundo se cierra la compuerta de inactivación y los iones sodio ya no pueden pasar hacia el interior de la membrana. En este punto el potencial de membrana comienza a recuperarse de nuevo hacia el estado de membrana en reposo, lo que es el proceso de repolarización. Otra característica importante de la inactivación del canal de sodio es que la compuerta de inactivación no se abre de nuevo hasta que el potencial de membrana se normaliza o casi a valores de reposo. Por tanto, en general el canal de sodio no se puede abrir de nuevo sin que antes se repolarice la fibra nerviosa.
Funciones de otros iones durante el potencial de acción
Iones con carga negativa (aniones) no difusibles en el interior del axón nervioso
En el interior del axón hay muchos iones de carga negativa que no pueden atravesar los canales de la membrana. Incluyen los aniones de las moléculas proteicas y de muchos compuestos de fosfato orgánicos, compuestos de sulfato y similares. Como estos iones no pueden salir del interior del axón, cualquier déficit de iones positivos en el interior de la membrana deja un exceso de estos aniones negativos no difusibles. Por tanto, estos iones negativos no difusibles son responsables de la carga negativa en el interior de la fibra cuando hay un déficit neto de iones potasio de carga positiva y deotros iones positivos.
Iones calcio
Las membranas de casi todas las células del cuerpo tienen una bomba de calcio similar a la bomba de sodio, y el calcio coopera con el sodio (o actúa en su lugar) en algunas células para producir la mayor parte del potencial de acción. Al igual que la bomba de sodio, la bomba de potasio transporta iones calcio desde el interior hacia el exterior de la membrana celular (o hacia el interior del retículo endoplásmico de la célula), creando un gradiente de ion calcio de aproximadamente 10.000 veces.
Inicio del potencial de acción
Un ciclo de retroalimentación positiva abre los canales de sodio
Este proceso es un círculo vicioso de retroalimentación positiva que, una vez que la retroalimentación es lo suficientemente intensa, continúa hasta que se han activado (abierto) todos los canales de sodio activados por el voltaje. Posteriormente, en un plazo de otra fracción de milisegundo, el aumento del potencial de membrana produce cierre de los canales de sodio y apertura de los canales de potasio, y pronto finaliza el potencial de acción.
Umbral para el inicio del potencial de acción
No se producirá un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen al ciclo de retroalimentación positiva que se ha descrito en el párrafo anterior. Esto se produce cuando el número de iones sodio que entran en la fibra supera al número de iones potasio que salen. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Por tanto, un aumento súbito del potencial de membrana en una fibra nerviosa grande desde –90 mV hasta aproximadamente –65 mV suele dar lugar a la aparición explosiva de un potencial de acción. Se dice que este nivel de –65 mV es el umbral para la estimulación.
La propagación de cada potencial de acción a lo largo de una fibra nerviosa reduce ligeramente las diferencias de concentración de sodio y de potasio en el interior y en el exterior de la membrana, porque los iones sodio difunden hacia el interior durante la despolarización y los iones potasio difunden hacia el exterior durante la repolarización. Para un único potencial de acción este efecto es tan pequeño que no se puede medir, con el tiempo se hace necesario restablecer las diferencias de las concentraciones de membrana de sodio y de potasio, que se consiguen por la acción de la bomba Na+-K+ de la misma manera que se ha descrito previamente para el restablecimiento original del potencial en reposo. Es decir, los iones sodio que han difundido hacia el interior de la célula durante los potenciales de acción y los iones potasio que han difundido hacia el exterior deben volver a su estado original por la bomba Na+-K+. Como esta bomba precisa energía para esta operación, esta «recarga» de la fibra nerviosa es un proceso metabólico activo que utiliza la energía que procede del sistema energético del trifosfato de adenosina de la célula.
En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y solo después comienza la repolarización, Este tipo de potencial de acción se produce en las fibras musculares cardíacas, en las que la meseta dura hasta 0,2 a 0,3 s y hace que la contracción del músculo cardíaco dure este mismo y prolongado período de tiempo.
La causa de la meseta es una combinación de varios factores. En primer lugar, en el proceso de despolarización del músculo cardíaco participan dos tipos de canales: 1) los canales de sodio habituales activados por el voltaje, denominados canales rápidos, y 2) los canales de calcio-sodio activados por el voltaje (canales de calcio de tipo L), que tienen una apertura lenta y que, por tanto, se denominan canales lentos.
Conducción «saltatoria» en las fibras mielinizadas de un nódulo a otro
Aunque los iones apenas pueden fluir a través de las gruesas vainas de mielina de los nervios mielinizados, sí lo hacen fácilmente a través de los nódulos de Ranvier. Por tanto, los potenciales de acción se producen solo en los nódulos. A pesar de todo, los potenciales de acción se conducen desde un nódulo a otro.
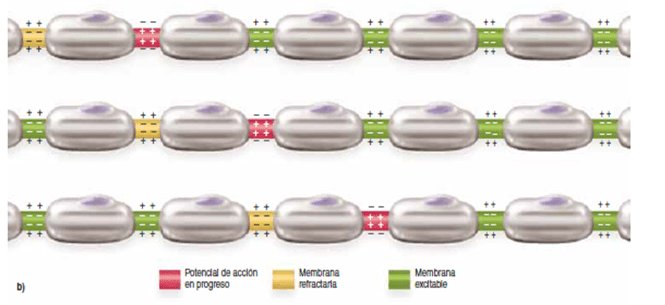
La conducción saltatoria es útil por dos motivos. Primero, al hacer que el proceso de despolarización salte intervalos largos a lo largo del eje de la fibra nerviosa, este mecanismo aumenta la velocidad de la transmisión nerviosa en las fibras mielinizadas hasta 5 a 50 veces. Segundo, la conducción saltatoria conserva la energía para el axón porque solo se despolarizan los nódulos, permitiendo una pérdida de iones tal vez 100 veces menor de lo que sería necesario de otra manera, y por tanto precisa poco gasto de energía para restablecer las diferencias de concentración de sodio y de potasio a través de la membrana después de una serie de impulsos nerviosos.
Excitación: el proceso de generación del potencial de acción
Esta apertura regenerativa automática se puede deber a un trastorno mecánico de la membrana, a los efectos químicos sobre la membrana o al paso de electricidad a través de la membrana. Todos estos enfoques se utilizan en diferentes puntos del cuerpo para generar potenciales de acción nerviosos o musculares: presión nerviosa para excitar las terminaciones nerviosas sensitivas de la piel, neurotransmisores químicos para transmitir señales desde una neurona a la siguiente en el cerebro y una corriente eléctrica para transmitir señales entre células musculares sucesivas del corazón y del intestino
Umbral de excitación y «potenciales locales agudos»
Un estímulo eléctrico negativo débil puede no ser capaz de excitar una fibra. Sin embargo, cuando aumenta el voltaje del estímulo se llega a un punto en el que se produce la excitación.
Inhibición de la excitabilidad: «estabilizadores» y anestésicos locales
Al contrario de los factores que aumentan la estabilidad nerviosa, otros factores, denominados factores estabilizadores de la membrana, pueden reducir la excitabilidad. Por ejemplo, una concentración elevada de calcio en el líquido extracelular reduce la permeabilidad de la membrana a los iones sodio y reduce simultáneamente la excitabilidad. Por tanto, se dice que el ion calcio es un «estabilizador»
«Período refractario» tras un potencial de acción, durante el cual no se puede generar un nuevo estímulo
No se puede producir un nuevo potencial de acción en una fibra excitable mientras la membrana siga despolarizada por el potencial de acción precedente. El motivo de esta restricción es que poco después del inicio del potencial de acción se inactivan los canales de sodio (o los canales de potasio, o ambos), y ninguna magnitud de la señal excitadora que se aplique a estos canales en este momento abrirá las compuertas de inactivación. La única situación que permitirá que se vuelvan a abrir es que el potencial de membrana vuelva al nivel del potencial de membrana en reposo original o cerca de este. Entonces, en otra pequeña fracción de segundo se abren las compuertas de inactivación del canal y se puede iniciar un nuevo potencial de acción. El período durante el cual no se puede generar un segundo potencial de acción, incluso con un estímulo intenso, se denomina período refractario absoluto.
😃
