LOS REFLEJOS MEDULARES

La información sensitiva se integra a todos los niveles del sistema nervioso y genera las respuestas motoras adecuadas que comienzan en la médula espinal con los reflejos musculares relativamente sencillos, se extienden hacia el tronco del encéfalo con unas actividades más complicadas y finalmente alcanzan el cerebro, donde están controladas las tareas musculares más complejas. Sin los circuitos neuronales especiales de la médula, hasta los sistemas de regulación motora más complejos del cerebro serían incapaces de causar cualquier movimiento muscular voluntario. En cambio, los circuitos encargados de estos movimientos están en la médula, y el cerebro no hace más que enviar señales que hacen llegar órdenes a la médula espinal para poner en acción el proceso de la marcha.
Organización de la médula espinal para las funciones motoras
La sustancia gris medular es la zona de integración para los reflejos medulares. Las señales sensitivas penetran en ella por las raíces sensitivas, también conocidas como raíces posteriores o dorsales.
Motoneuronas anteriores
En ellas nacen las fibras nerviosas que salen de la médula a través de las raíces anteriores e inervan directamente las fibras de los músculos esqueléticos. Estas neuronas son de dos tipos, motoneuronas α y motoneuronas γ.
Motoneuronas γ
son mucho más pequeñas que están situadas en las astas anteriores de la médula espinal. Estas células transmiten impulsos a través de unas fibras nerviosas motoras γ de tipo A (Aγ) mucho más pequeñas, con un diámetro
medio de 5 μm, que van dirigidas hacia unas fibras del músculo esquelético especiales pequeñas llamadas fibras intrafusales. Estas fibras ocupan el centro del huso muscular, que sirve para controlar el «tono» básico del músculo.
Interneuronas
Están presentes en todas las regiones de la sustancia gris medular.
Su tamaño es pequeño y poseen una naturaleza muy excitable, muestran una actividad espontánea capaz de emitir hasta 1.500 disparos por segundo. Entre sí presentan múltiples interconexiones y muchas de ellas también establecen sinapsis directas con las motoneuronas anteriores.
Receptores sensitivos musculares (husos musculares y órganos tendinosos de Golgi) y sus funciones en el control muscular
El control adecuado del funcionamiento muscular exige la excitación del músculo por parte de las motoneuronas anteriores de la médula espinal y también una retroalimentación permanente con la información sensitiva que llega a ella procedente de cualquier músculo, para indicar su estado
funcional en cada momento. Para comunicar esta información, los músculos y sus tendones reciben una inervación abundante por parte de dos tipos especiales de receptores sensitivos:
- los husos musculares, que están distribuidos por todo el vientre muscular y envían información hacia el sistema nervioso sobre la longitud del músculo o la velocidad con la que varía esta magnitud
- los órganos tendinosos de Golgi que se encuentran situados en los tendones musculares y transmiten información sobre la tensión tendinosa o su ritmo de cambio. Las señales procedentes de estos dos receptores tienen como propósito casi exclusivo el control muscular intrínseco.
- Función receptora del huso muscular: Estructura e inervación motora del huso muscular. Cada elemento tiene una longitud de 3 a 10 mm. Se encuentra dispuesto alrededor de 3 a 12 fibras musculares intrafusales diminutas cuyos extremos acaban en punta y se fijan al glucocáliz de las grandes fibras extrafusales adyacentes correspondientes al músculo esquelético. Cualquier fibra muscular intrafusal es una fibra muscular esquelética muy pequeña. Sin embargo, su región central, es decir, el área equidistante entre sus dos extremos, contiene pocos filamentos de actina y miosina o ninguno. Esta parte central no se contrae cuando lo hacen sus extremos.Funciona como un receptor sensitivo, Las porciones finales que sí se contraen reciben su excitación de fibras nerviosas motoras γ de tamaño reducido que nacen en las pequeñas motoneuronas γ de tipo A situadas en las astas anteriores de la médula espinal, Estas fibras nerviosas motoras γ también se denominan fibras eferentes γ, en contraposición a las fibras eferentes α grandes (fibras nerviosas α de tipo A) que inervan el músculo esquelético extrafusal.
- Inervación sensitiva del huso muscular: La porción receptora del huso muscular se localiza en su parte central. En esta zona, las fibras musculares intrafusales carecen de los elementos contráctiles miosina y actina. Es fácil comprobar que el receptor del huso muscular puede excitarse por dos mecanismos:El alargamiento del músculo en su conjunto estira la porción intermedia del huso y, por tanto, estimula al receptor.
aunque la longitud de todo el musculo no cambie, la contraccion de las porciones finales de las fibras intrafusales tambien estira la porcion intermedia del huso y asi activa el receptor
Terminación primaria
En el centro de la zona receptora, una gran fibra nerviosa sensitiva rodea la porción central de cada fibra intrafusal, formando la denominada terminación aferente primaria o terminación anuloespiral.
Respuesta de las terminaciones primarias y secundarias a la longitud del receptor: respuesta «estática»
Este efecto se llama respuesta estática del receptor del huso, lo que quiere decir que las terminaciones primarias y secundarias siguen enviando sus impulsos durante varios minutos como mínimo si el propio huso muscular permanece estirado.
Respuesta de la terminación primaria a la velocidad de cambio en la longitud del receptor: respuesta «dinámica»
Cuando la longitud del receptor del huso aumenta de forma repentina, la terminación primaria recibe un estímulo potente. Este estímulo se denomina respuesta dinámica, lo que significa que la terminación primaria responde de un modo vivísimo a una velocidad de cambio rápida en la longitud del huso. Por tanto, la terminación primaria manda unos impulsos potentísimos hacia la médula espinal, positivos o negativos, para comunicar cualquier cambio ocurrido en la longitud del receptor del huso.
Reflejo miotático muscular
La manifestación más sencilla del funcionamiento del huso es el reflejo miotático o de estiramiento muscular. Siempre que se estira bruscamente un músculo, la activación de los husos causa la contracción refleja de las fibras musculares esqueléticas grandes en el músculo estirado y también en
los músculos sinérgicos más íntimamente ligados.
Circuito neuronal del reflejo miotático

En él aparece una fibra nerviosa propioceptora de tipo Ia que se origina en un huso muscular y penetra por una raíz posterior de la médula espinal. luego una rama de esta fibra se encamina directamente hacia
el asta anterior de la sustancia gris medular y hace sinapsis con las motoneuronas anteriores que devuelven fibras nerviosas motoras al mismo músculo en el que se había originado la fibra del huso. Esta vía monosináptica permite el regreso al músculo de una señal refleja en el menor lapso de tiempo posible después de la excitación del huso. La mayoría de las fibras de tipo II procedentes del huso muscular acaban en numerosas interneuronas de la sustancia gris medular, que a su vez transmiten impulsos retardados hacia las motoneuronas anteriores o cumplen otras funciones.
Reflejos miotáticos dinámico y estático
El reflejo miotático puede dividirse en dos componentes: el dinámico y el estático.
El reflejo miotático dinámico surge con potentes señales dinámicas transmitidas desde las terminaciones sensitivas primarias de los husos musculares, originada por su estiramiento o distensión rápida. Esto pasa cuando un músculo se estira o se distiende bruscamente, se transmite un impulso potente hacia la médula espinal, lo que provoca instantáneamente una enérgica contracción refleja (o un descenso de la contracción) en el mismo músculo del que nació la señal.
El reflejo sirve para oponerse a los cambios súbitos sufridos en la longitud muscular.
El reflejo miotático dinámico finaliza una fracción de segundo después de que el músculo se haya estirado (o distendido) hasta alcanzar su nueva longitud, pero después le sigue un reflejo miotático estático más débil que se mantiene un período prolongado desde ese instante. Este reflejo deriva de
las señales receptoras estáticas continuas transmitidas por las terminaciones primarias y secundarias.
La importancia del reflejo miotático estático radica en que produce un grado de contracción muscular que puede mantenerse razonablemente constante.
Función «amortiguadora» de los reflejos miotáticos dinámico y estático en la
contracción del músculo liso
Una misión especialmente importante del reflejo miotático es su capacidad para evitar las oscilaciones o las sacudidas en los movimientos corporales, que es una función amortiguadora o suavizadora.
Los impulsos de la médula espinal muchas veces se transmiten hasta un músculo según un patrón irregular, con un aumento de su intensidad que dura unos pocos milisegundos y después un descenso, Cuando el aparato del huso muscular no funciona satisfactoriamente, la contracción del músculo adquiere un carácter entrecortado durante el curso de dicha señal.
Intervención del huso muscular en la actividad motora voluntaria
El 31% de todas las fibras nerviosas motoras dirigidas al músculo son fibras eferentes de tipo A pequeñas en vez de las fibras motoras de tipo A grandes. Este efecto hace que se contraigan al mismo tiempo las fibras musculares esqueléticas extrafusales y las fibras intrafusales del huso muscular. El objetivo de la contracción simultánea de las fibras intrafusales del huso muscular y de las grandes fibras musculares esqueléticas es doble. Por ejemplo, si el huso muscular no se contrajera y relajara al unísono con las grandes fibras musculares, a veces su porción receptora estaría oscilando y otras veces se encontraría hiperestirada, sin que en ninguno de estos casos operase dentro de las condiciones óptimas para cumplir su función.
dado que la región facilitadora bulborreticular está especialmente relacionada con las contracciones antigravitatorias, y que los músculos antigravitatorios poseen una densidad especialmente alta de husos musculares, se insiste en la importancia del mecanismo eferente para amortiguar los movimientos de las diversas partes del cuerpo durante la marcha y la carrera.
El sistema de los husos musculares estabiliza la posición corporal durante una acción a tensión
Una de las funciones más importantes que desempeña el sistema de los husos musculares consiste en estabilizar la posición corporal durante las acciones motoras a tensión. Para realizar esta función, la región facilitadora bulborreticular y sus zonas afines del tronco del encéfalo transmiten señales estimuladoras hacia las fibras musculares intrafusales del huso muscular a través de las fibras nerviosas . Esta acción acorta los extremos del huso y estira sus regiones receptoras centrales, lo que aumenta la frecuencia de emisión de sus impulsos. En cualquier momento en que una persona tenga que ejecutar una función muscular que exija una postura muy delicada y exacta, la excitación de los husos musculares adecuados por parte de las señales procedentes de la región facilitadora bulborreticular del tronco del encéfalo estabiliza la posición de la mayoría de las articulaciones
Reflejo tendinoso de Golgi
El órgano tendinoso de Golgi sirve para controlar la tensión muscular
El órgano tendinoso de Golgi es un receptor sensitivo encapsulado
por el que pasan las fibras del tendón muscular. Cada órgano tendinoso de Golgi suele estar conectado con unas 10 a 15 fibras musculares, que lo estimulan cuando este pequeño haz se «tensa» debido a la contracción o el estiramiento del músculo. Por tanto, la principal diferencia en la excitación del órgano tendinoso de Golgi en comparación con el huso muscular reside en que el huso detecta la longitud del músculo y los cambios de la misma, mientras que el órgano tendinoso identifica la tensión muscular, según queda patente por su propio grado.
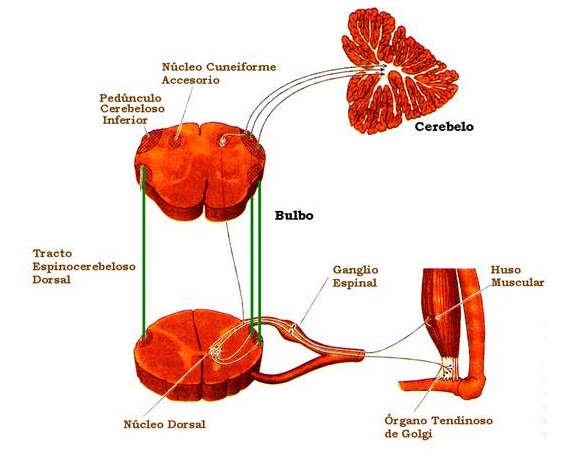
El órgano tendinoso, lo mismo que el receptor primario del huso muscular, ofrece una respuesta dinámica y una respuesta estática
Reflejo flexor y reflejos de retirada
En el animal espinal o descerebrado es fácil que prácticamente cualquier tipo de estímulo sensitivo cutáneo de los miembros haga que sus músculos flexores se contraigan, lo que permite retirar la extremidad del objeto estimulador. Es el denominado reflejo flexor.
En su forma clásica, el reflejo flexor se suscita con mayor potencia mediante la estimulación de las terminaciones para el dolor, como sucede con un pinchazo, el calor o una herida, razón por la que también se le denomina reflejo nociceptivo. La activación de los receptores para el tacto también puede despertar un reflejo flexor más débil y menos prolongado.
Si cualquier parte del cuerpo aparte de las extremidades recibe un estímulo doloroso, esa porción se alejará del estímulo en correspondencia, pero el reflejo puede no quedar limitado a los músculos flexores, aun cuando sea básicamente el mismo tipo de fenómeno. Por tanto, cualquiera de los
múltiples patrones que adoptan en las diferentes regiones del organismo se llama reflejo de retirada.

Marcha recíproca de las extremidades opuestas
Si la médula espinal lumbar no se secciona hasta el centro, cada vez que se den unos pasos en sentido hacia delante con una extremidad, la opuesta corrientemente se desplaza hacia atrás. Este efecto deriva de la inervación recíproca existente entre ambos miembros. Marcha en diagonal entre las cuatro extremidades: el reflejo de «marcar el paso»
En general, los pasos siguen un patrón en diagonal entre las patas
delanteras y las traseras. Esta respuesta diagonal constituye otra manifestación de la inervación recíproca, esta vez a lo largo de toda la longitud de la médula hacia arriba y hacia abajo entre las
extremidades anteriores y las posteriores. Este patrón de marcha se denomina reflejo de marcar el paso.
Reflejo de galope
en el que las extremidades anteriores se desplazan hacia atrás al unísono a la vez que las posteriores se mueven hacia delante. Este reflejo suele suceder cuando se aplican estímulos casi idénticos de estiramiento o de presión a las extremidades de ambos lados del cuerpo al mismo tiempo: su estimulación dispar promueve el reflejo de la marcha en diagonal. Esto encaja con los patrones normales de la marcha y el galope, porque al caminar, cada vez no se estimula nada más que una pata delantera y otra trasera, lo que pondría al animal en condiciones de seguir avanzando. En cambio, al golpear el suelo durante el galope, las dos extremidades anteriores y las dos posteriores se estimulan más o menos por igual, lo que le deja listo para continuar galopando y, por tanto, mantener este patrón de movimiento.
