El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto porque la duración de la contracción es mucho mayor.

el músculo cardíaco tiene las miofibrillas típicas que contienen filamentos de
actina y de miosina casi idénticos a los que se encuentran en el músculo esquelético; estos filamentos están unos al lado de otros y se deslizan durante la contracción de la misma manera que ocurre en el músculo esquelético.
músculo cardíaco es un sincitio:
Las zonas oscuras que atraviesan las fibras musculares cardíacas se le denominan
discos intercalados, las fibras musculares cardíacas están formadas por muchas células
individuales conectadas entre sí en serie y en paralelo.

En cada uno de los discos intercalados las membranas celulares se fusionan entre sí para formar uniones «comunicantes» (en hendidura) permeables que permiten una rápida difusión.
desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular a lo largo del eje longitudinal de las fibras musculares cardíacas, de modo que los potenciales de acción viajan fácilmente desde una célula muscular cardíaca a la siguiente, a través de los discos intercalados. Por tanto, el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una célula se excita el potencial de
acción se propaga rápidamente a todas.
El corazón realmente está formado por dos sincitios: el sincitio auricular, que forma las paredes de las dos aurículas, y el sincitio ventricular, que forma las paredes de los dos ventrículos. Las aurículas están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares (AV) entre las aurículas y los ventrículos. Normalmente los potenciales no se conducen desde el sincitio auricular hacia el sincitio ventricular directamente a través de este tejido fibroso. Por el contrario, solo son conducidos por medio de un sistema de conducción especializado
denominado haz AV, que es un fascículo de fibras de conducción de varios milímetros de diámetro.
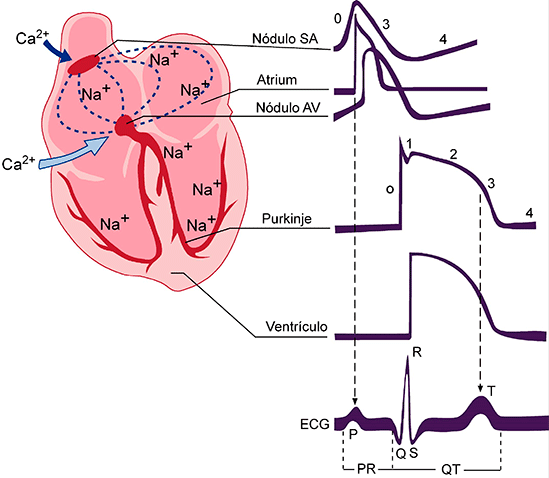
Potenciales de acción en el músculo cardíaco:
El potencial de acción que se registra en una fibra muscular ventricular, es en promedio de aproximadamente 105 mV, lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproximadamente –85 mV, entre los latidos
hasta un valor ligeramente positivo, de aproximadamente +20 mV, durante cada latido. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta, seguida al final de la meseta de una repolarización súbita. La presencia de esta meseta del potencial de acción ha- ce que la contracción ventricular dure hasta 15 veces más en el músculo cardíaco que en el músculo esquelético.
el potencial de acción del músculo esquelético está producido casi por completo
por la apertura súbita de grandes números de canales rápidos de sodio que permiten que grandes cantidades de iones sodio entren en la fibra muscular esquelética desde el líquido extracelular. A estos canales se les denomina canales «rápidos» porque permanecen abiertos solo algunas milésimas de segundo y después se cierran súbitamente. Al final de este cierre se produce la repolarización y el potencial de acción ha terminado en otra milésima de segundo aproximadamente
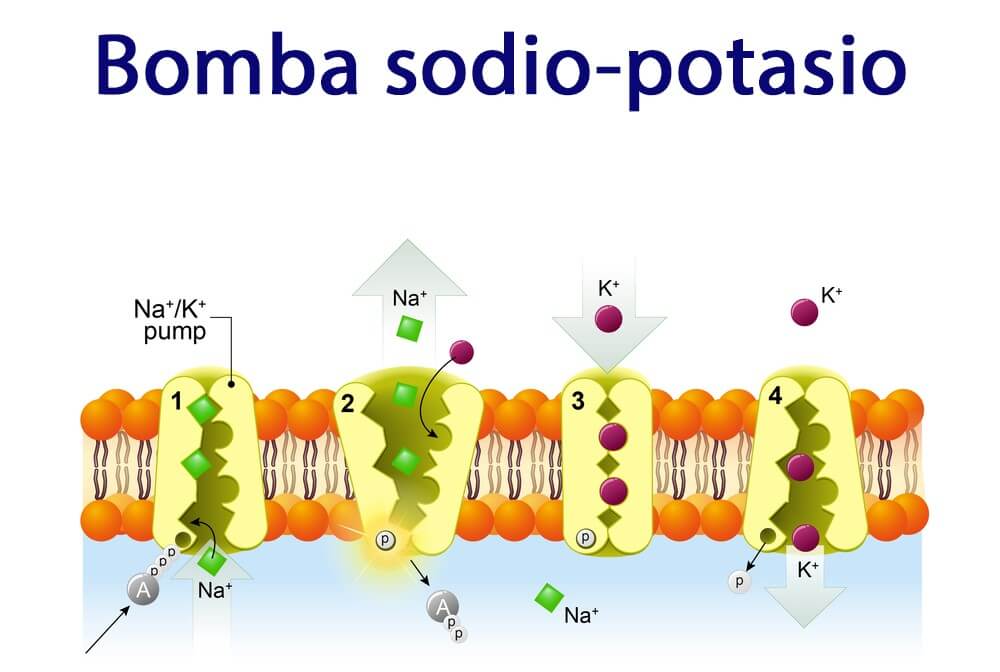
el potencial de acción está producido por la apertura de dos tipos de
canales: 1) los mismos canales rápidos de sodio activados por el voltaje que en el músculo.
esquelético y 2) otra población totalmente distinta de canales de calcio de tipo L (canales lentos de calcio), que también se denominan canales de calcio-sodio.
La segunda diferencia funcional importante entre el músculo cardíaco y el músculo esquelético que ayuda a explicar tanto el potencial de acción prolongado como su meseta es la siguiente: inmediatamente después del inicio del potencial de acción, la permeabilidad de la membrana del músculo cardíaco a los iones potasio disminuye aproximadamente cinco veces, un efecto que no aparece en el músculo esquelético.
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de unos 0,3 a 0,5 m/s, o aproximadamente 1/250 de la velocidad en las fibras nerviosas grandes y en torno a 1/10 de la velocidad en las fibras musculares esqueléticas. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje, es de hasta 4 m/s en la mayoría de las partes del sistema, lo que permite una conducción razonablemente rápida de la señal excitadora hacia las diferentes partes del corazón.

Duración de la contracción
El músculo cardíaco comienza a contraerse algunos milisegundos después de la llegada del potencial de acción y sigue contrayéndose hasta algunos milisegundos después de que finalice. la duración de la contracción del músculo cardíaco depende principalmente de la duración del potencial de acción, incluyendo la meseta, aproximadamente 0,2 s en el músculo auricular y 0,3 s en el músculo ventricular.
Diástole y sístole
El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole.
La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca. Por ejemplo, si la frecuencia cardíaca es de 72 latidos/min, la duración del ciclo cardíaco es de 1/72 min/latido, aproximadamente 0,0139 min por latido, o 0,833 s por latido.
los diferentes acontecimientos que se producen durante el ciclo cardíaco
para el lado izquierdo del corazón. Las tres curvas superiores muestran los cambios de presión en la aorta, en el ventrículo izquierdo y en la aurícula izquierda, respectivamente. La cuarta curva representa los cambios del volumen ventricular izquierdo, la quinta el electrocardiograma y la sexta un fonocardiograma, que es un registro de los ruidos que produce el corazón (principalmente las
válvulas cardíacas) durante su función de bombeo.
La sangre normalmente fluye de forma continua desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas. Después, la contracción auricular habitualmente produce un llenado de un 20% adicional de los ventrículos. Por tanto, las aurículas actúan como bombas de cebado que aumentan la eficacia del bombeo ventricular hasta un 20%. el corazón puede seguir funcionando en la mayor parte de las condiciones incluso sin esta
eficacia de un 20% adicional porque normalmente tiene la capacidad de bombear entre el 300 y el 400% más de sangre de la que necesita el cuerpo en reposo. Por tanto, cuando las aurículas dejan de funcionar es poco probable que se observe esta diferencia salvo que la persona haga un esfuerzo; en este caso de manera ocasional aparecen síntomas agudos de insuficiencia cardíaca, especialmente disnea.
Los ventrículos se llenan de sangre durante la diástole:
Durante la sístole ventricular se acumulan grandes cantidades de sangre en las aurículas derecha e izquierda porque las válvulas AV están cerradas. Por tanto, tan pronto como ha finalizado la sístole y las presiones ventriculares disminuyen de nuevo a sus valores diastólicos bajos, el aumento moderado de presión que se ha generado en las aurículas durante la sístole ventricular
inmediatamente abre las válvulas AV y permite que la sangre fluya rápidamente hacia los ventrículos, como se muestra en la elevación de la curva de volumen ventricular izquierdo. Es el
denominado período de llenado rápido de los ventrículos.
El período de llenado rápido dura aproximadamente el primer tercio de la diástole. Durante el tercio medio de la diástole normalmente solo fluye una pequeña cantidad de sangre hacia los ventrículos; esta es la sangre que continúa drenando hacia las aurículas desde las venas y que pasa a través de las aurículas directamente hacia los ventrículos.
Durante el último tercio de la diástole las aurículas se contraen y aportan un impulso adicional al flujo de entrada de sangre hacia los ventrículos.

Válvulas auriculoventriculares
Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmona hacia los ventrículos durante la diástole. para el
ventrículo izquierdo, se cierran y abren pasivamente. Es decir, se cierran cuando un gradiente de presión retrógrada empuja la sangre hacia atrás, y se abren cuando un gradiente de presión anterógrada fuerza la sangre en dirección anterógrada. Por motivos anatómicos, las válvulas AV, que están formadas por una película delgada, casi no precisan ningún flujo retrógrado para cerrarse, mientras que las válvulas semilunares, que son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
Los músculos papilares
se contraen cuando se contraen las paredes
ventriculares, pero, al contrario de lo que se podría esperar, no contribuyen al cierre de las válvulas. Por el contrario, tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular. Si se produce la rotura de una cuerda tendinosa o si se produce parálisis de uno de los músculos papilares, la válvula protruye mucho hacia las aurículas durante la contracción ventricular, a veces tanto que se produce una fuga grave y da lugar a una insuficiencia cardíaca grave o incluso mortal.
Válvulas aórtica y de la arteria pulmonar
Las válvulas semilunares aórtica y pulmonar funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente, a diferencia del cierre mucho más suave de las válvulas AV. Segundo, debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores. Además, debido al cierre rápido y a la eyección rápida, los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV. Las válvulas AV tienen el soporte de las cuerdas tendinosas, lo que no ocurre en el caso de las válvulas semilunares. A partir de la anatomía de las válvulas aórtica y pulmonar (que se muestra para la válvula aórtica en la parte inferior de la figura 9-8) es evidente que deben estar situadas sobre una base de un tejido fibroso especialmente fuerte, pero muy flexible para soportar las tensiones físicas adicionales.
Válvulas aórtica y de la arteria pulmonar
Las válvulas semilunares aórtica y pulmonar
funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente, a diferencia del cierre mucho más suave de las válvulas AV
Segundo, debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores. Además, debido al cierre rápido y a la eyección rápida, los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV. Las válvulas AV tienen el soporte de las cuerdas tendinosas, lo que no ocurre en el caso de las válvulas
semilunares. A partir de la anatomía de las válvulas aórtica y pulmonar (que se muestra para la válvula aórtica en la parte inferior, es evidente que deben estar situadas sobre una base de un tejido fibroso especialmente fuerte, pero muy flexible para soportar las tensiones físicas adicionales.

Regulación del bombeo cardíaco
Cuando una persona está en reposo el corazón solo bombea de 4 a 6 l de sangre cada minuto. Durante el ejercicio intenso puede ser necesario que el corazón bombee de cuatro a siete veces esta cantidad.
Los mecanismos básicos mediante los que se regula el volumen que bombea el corazón son:
1)regulación cardíaca intrínseca del bombeo en respuesta a los cambios del volumen de la sangre que
fluye hacia el corazón
2) control de la frecuencia cardíaca y del bombeo cardíaco por el sistema
nervioso autónomo.
Mecanismo de Frank-Starling:
La capacidad intrínseca del corazón de adaptarse a volúmenes crecientes de flujo sanguíneo de entrada se denomina mecanismo de Frank-Starling del corazón en honor de Otto Frank y Ernest Starling, dos grandes fisiólogos de hace un siglo. Básicamente, el mecanismo de Frank-Starling significa que cuanto más se distiende el músculo cardíaco durante el llenado, mayor es la fuerza de contracción y mayor es la cantidad de sangre que bombea hacia la aorta. O, enunciado de otra manera, dentro de límites fisiológicos el corazón bombea toda la sangre que le llega procedente de las venas.
Cuando una cantidad adicional de sangre fluye hacia los ventrículos, el propio músculo cardíaco es distendido hasta una mayor longitud. Esta distensión, a su vez, hace que el músculo se contraiga con más fuerza porque los filamentos de actina y de miosina son desplazados hacia un grado más óptimo de superposición para la generación de fuerza. Por tanto, el ventrículo, debido al aumento de la función de bomba, bombea automáticamente la sangre adicional hacia las arterias. Esta capacidad del músculo distendido, hasta una longitud óptima, de contraerse con un aumento
del trabajo cardíaco, es característica de todo el músculo estriado.
no es simplemente una característica del músculo cardíaco. Es importante efecto del aumento de longitud del músculo cardíaco, hay otro factor que aumenta la función de bomba del corazón cuando aumenta su volumen. La distensión de la pared de la aurícula derecha aumenta directamente la frecuencia cardíaca en un 10-20%, lo que también contribuye a aumentar la cantidad de sangre que se bombea cada minuto, aunque su contribución es mucho menor que la del mecanismo de Frank-Starling.
Control del corazón por los nervios simpáticos y parasimpáticos
La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos y parasimpáticos (vagos), que inervan de forma abundante el corazón.
Para niveles dados de presión auricular de entrada, la cantidad de sangre que se bombea cada minuto (gasto cardíaco) con frecuencia se puede aumentar más de un 100% por la estimulación simpática. Por el contrario, el gasto se puede disminuir hasta un valor tan bajo como cero o casi cero por la estimulación vagal (parasimpática).
La estimulación simpática intensa puede aumentar la frecuencia cardíaca en seres humanos adultos jóvenes desde la frecuencia normal de 70 latidos/min hasta 180 a 200 y, raras veces, incluso 250 latidos/min. Además, la estimulación simpática aumenta la fuerza de la contracción cardíaca hasta el doble de lo normal, aumentando de esta manera el volumen de sangre que se bombea y aumentando la presión de eyección. Así, con frecuencia la estimulación simpática puede aumentar el gasto cardíaco máximo hasta dos o tres veces, además del aumento del gasto que produce el mecanismo de Frank-Starling que ya se ha comentado. Por el contrario, la inhibición de los nervios simpáticos del corazón puede disminuir la función de bomba del corazón en un grado moderado: en condiciones normales, las fibras nerviosas simpáticas que llegan al corazón descargan continuamente a una frecuencia baja que mantiene el bombeo aproximadamente un 30% por encima del que habría sin estimulación simpática. Por tanto, cuando la actividad del sistema nervioso simpático disminuye por debajo de lo normal, tanto de la frecuencia cardíaca como la fuerza de la contracción del músculo ventricular se reducen, con lo que disminuye el nivel de bombeo cardíaco hasta un 30% por debajo de lo normal.
La estimulación parasimpática (vagal) reduce la frecuencia cardíaca y la fuerza de la contracción
La estimulación intensa de las fibras nerviosas parasimpáticas de los nervios vagos que llegan al corazón puede interrumpir el latido cardíaco durante algunos segundos, pero después el corazón habitualmente «escapa» y late a una frecuencia de 20 a 40 latidos/min mientras continúe la estimulación parasimpática. Además, la estimulación vagal intensa puede reducir la fuerza de la contracción del músculo cardíaco en un 20-30%.
Las fibras vagales se distribuyen principalmente por las aurículas y no mucho en los ventrículos, en los que se produce la contracción de potencia del corazón. Esta distribución explica por qué el efecto de la estimulación vagal tiene lugar principalmente sobre la reducción de la frecuencia cardíaca, en lugar de reducir mucho la fuerza de la contracción del corazón. Sin embargo, la gran disminución de la frecuencia cardíaca, combinada con una ligera reducción de la fuerza de la contracción cardíaca, puede reducir el bombeo ventricular en un 50% o más.
Efecto de los iones potasio
El exceso de potasio hace que el corazón esté dilatado y flácido, y también reduce la frecuencia cardíaca. Grandes cantidades de potasio también pueden bloquear la conducción del impulso cardíaco desde las aurículas hacia los ventrículos a través del haz AV. La elevación de la concentración de potasio hasta solo 8 a 12 mEq/l (dos a tres veces el valor normal) puede producir una profunda debilidad del corazón, una alteración del ritmo e incluso la muerte.
Estos efectos se deben parcialmente al hecho de que una concentración elevada de potasio en los líquidos extracelulares reduce el potencial de membrana en reposo de las fibras del músculo cardíaco, como se explica en el capítulo 5. Es decir, la alta concentración de potasio en el líquido extracelular despolariza parcialmente la membrana celular, lo que provoca que el potencial de membrana sea menos negativo. Cuando disminuye el potencial de membrana también lo hace la intensidad del potencial de acción, lo que hace que la contracción del corazón sea progresivamente
más débil.
Efecto de los iones calcio
Un exceso de iones calcio produce efectos casi exactamente contrarios a los de los iones potasio, haciendo que el corazón progrese hacia una contracción espástica. Este efecto está producido por el efecto directo de los iones calcio en el inicio del proceso contráctil cardíaco, como se explicó antes en este mismo capítulo.
Por el contrario, el déficit de iones calcio produce debilidad cardíaca, similar al efecto de la elevación de la concentración de potasio. Afortunadamente las concentraciones de iones calcio en la sangre normalmente están reguladas en un intervalo muy estrecho. Por tanto, los efectos cardíacos de las concentraciones anormales de calcio raras veces tienen significado clínico.
Efecto de la temperatura sobre la función cardíaca
El aumento de la temperatura corporal, como ocurre durante la fiebre, produce un gran aumento de la frecuencia cardíaca, a veces hasta del doble del valor normal. El descenso de la temperatura produce una gran disminución de la frecuencia cardíaca, que puede disminuir hasta solo algunos latidos por minuto cuando una persona está cerca de la muerte por hipotermia en el intervalo de temperatura corporal de 16 °C a 21 °C. Estos efectos probablemente se deben al hecho de que el calor
aumenta la permeabilidad de la membrana del músculo cardíaco a los iones que controlan la frecuencia cardíaca, acelerando el proceso de autoexcitación.
La fuerza contráctil del corazón con frecuencia se incrementa transitoriamente cuando hay un aumento moderado de la temperatura, como ocurre durante el ejercicio corporal, aunque una elevación prolongada de la temperatura agota los sistemas metabólicos del corazón y finalmente produce debilidad. Por tanto, la función óptima del corazón depende mucho del control adecuado de la temperatura corporal mediante los mecanismos