Organización funcional del cuerpo humano y control del medio interno
Para hablar de fisiología tenemos que empezar definiendo la.
FISIOLOGÍA: Es la ciencia que pretende explicar los mecanismos físicos y químicos responsables del origen, desarrollo y progresión de la vida.
Como ya sabemos cada tipo de vida por mas pequeño que sea posee sus propias características funcionales, por lo que la fisiología puede separarse en:
Fisiología Vírica
Fisiología bacteriana
Fisiología Celular
Fisiología Vegetal
Fisiología de los invertebrados
etc.
Pero por ahora nos enfocaremos en la fisiología humana.
FISIOLOGÍA HUMANA: Intenta explicar las características y mecanismos específicos del cuerpo humana que hacen que sea un ser vivo.
Las células como unidades vivas del cuerpo
La célula es la unidad viva básica del cuerpo.
cada órgano es un conjunto de ellas que se mantienen unidas mediante estructuras de soporte intercelulares. Esta especialmente diseñada para realizar una o mas funciones en concreto; ademas todas ellas tiene la capacidad de reproducirse formando mas células de su propia estirpe.
Liquido extracelular: el medio interno
El 60% del cuerpo humano es agua del adulto es liquido. casi todo este liquido se encuentra dentro de las células (liquido intracelular, LIC) una tercera parte se encuentra en los espacios exteriores de las células (liquido extracelular, LEC)
liquido extracelular
liquido intracelular
Iones sodio y cloruro, nutrientes Medio interno del organismo Milieu interieur –Fisiologo Frances Claude Bernaude Dióxido de carbono Movimiento constante por todo el cuerpo en dos etapas 1)Movimiento de la sangre dentro de los vasos sanguíneos 2)Movimiento entre los capilares sanguíneos y los espacios intercelulares
Iones potasio, magnesio y fosfato
HOMEOSTASIS
En 1929 el fisiologo estadounidense Walter Cannon acuño el termino homeostasis para referirse al mantenimiento de unas condiciones casi constantes del medio interno.
a veces se considera a la enfermedad como un estado de ruptura de la homeostasis, pero se sabe que incluso en presencia de la enfermedad, los mecanismos homeostaticos siguen activos y mantienen las funciones vitales.
La Fisiopatología: pretende explicar como se alteran los diversos procesos fisiológicos durante las enfermedades y las lesiones.
Los órganos y tejidos realizan diversas funciones que colaboran en el mantenimiento de estas condiciones relativamente constantes, por lo que hablaremos de alguno de ellos y como contribuyen a la homeostasis:
Aparato Respiratorio
Origen de los nutrientes en el LEC
Eliminación de los productos finales metabólicos
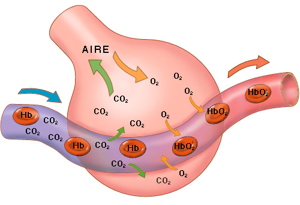
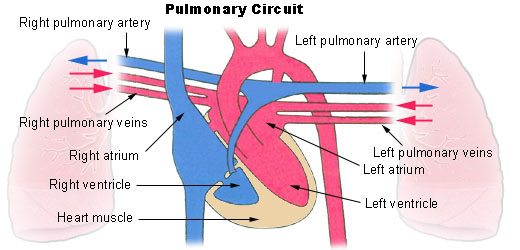
Cada vez que la sangre atraviesa el organismo también fluye por los pulmones y capta el oxigeno de los alvéolos, adquiriendo el oxigeno que necesitan las células. La membrana alveolar tiene un grosor de 0.4 a 2.0 µm lo que permite la difusión rápida del oxigeno
Al mismo tiempo que se capta el oxigeno, se libera el dióxido de carbono desde la sangre hacia los alvéolos y el movimiento respiratorio expulsa el dióxido de carbono del cuerpo
Aparato Digestivo
Origen de los nutrientes en el LEC
Eliminación de los productos finales metabólicos
cuando la sangre atraviesa las paredes del aparato digestivo absorbe los nutrientes (hidratos de carbono, acidos digestivos, y aa) y estos se llevan al LEC
Lo que ingerimos y no es digerido dentro de nuestro organismo es eliminado en las heces
Higado
Origen de los nutrientes en el LEC
Eliminación de los productos finales metabólicos
Es el encargado de cambiar la composición química de las sustancias, para convertirlas en formas mas utilizables
Es el encargado de La detoxificacion o eliminación de fármacos y productos químicos que ingerimos; Secreta estos residuos en la bilis para luego ser eliminadas en las heces
Aparato Locomotor
Los músculos contribuyen a la homeostasis, porque sin ellos el organismo no podría desplazarse para obtener los alimentos que se necesitan para la nutrición, también permite la movilidad como protección frente al entorno
Riñones
Con el paso de la sangre a través de los riñones se eliminan del plasma la mayoría de sustancias que las células ya no necesitan y también se eliminan los productos finales del metabolismo celular (urea, ácido úrico, exceso de iones y agua)
Regulación de las funciones corporales
El sistema nervioso se divide en:
La porción de aferencia sensitiva
El sistema nervioso central (o porción integradora)
Porción eferente motora
Los receptores sensitivos detectan el estado del cuerpo o su entorno. (La piel, ojos. oídos) estos son enviados al sistema nervioso central que esta formado por cerebro y médula espinal. El cerebro almacena la información, genera los pensamientos, crea la ambición y determina las reacciones que debe manifestar el cuerpo en repuesta a las sensaciones y se transmiten las señales a la porción motora eferente para llevar acabo los deseos del sujeto.
SISTEMA NERVIOSO AUTONOMO/NEUROVEGETATIVO
Funciona a una escala subconsciente y controla los funciones de los órganos internos
El Sistema Hormonal esta conformado por ocho glándulas endocrinas mayores y varios órganos y tejidos que segregan Hormonas.
Las hormas se transportan en el liquido extracelular a otras partes del cuerpo para regular las funciones celulares, las hormonas proporcionan un sistema de regulacion que complementa al sistema nervioso.
Protección del cuerpo
Sistema inmunitario
Esta formado por por glóbulos blancos, el timo, los nódulos linfáticos y los vasos linfáticos. Estos protegen el cuerpo de patógenos como bacterias, virus, parásitos y hongos.
Proporciona un mecanismo para que el cuerpo
Diferencia sus propias células de las células y sustancias extrañas
Destruyan al invasor por medio de fagocitosis o producción de linfocitos.
Sistema Tegumentario
Este sistema essta constituido por: La piel, el pelo, las uñas, las glandulas
Cubren, amortiguan y protegen los tejidos profundos y los órganos del cuerpo. La piel es nuestra primera barrera contra patógenos y separa el medio interno y el mundo exterior. también nos ayuda a la regulación de la temperatura y la excreción de los residuos (sudor). La piel suele comprender aprox entre el 12 y 15% del peso corporal.
Reproducción
Puede sonar un poco cruel dicho de esta forma pero la reproducción nos ayuda a mantener la homesotasis generando nuevos seres que ocuparan el lugar de aquellos que mueren.
Sistema de control del organismo.
Estos sistemas actúan dentro del organismo para controlar las funciones de sus componentes o las interrelaciones entre los órganos.
Ejemplo: existe un mecanismo encargado de mantener una concentración casi exacta y contante de oxigeno en el liquio extracelular. Como el oxigeno es una de las principales sustancias que requieren las reacciones químicas de las células. Este mecanismo funciona dependiendo de las características químicas de la hemoglobina, que esta presente en todos los eritrocitos. La hemoglobina se combina con el oxigeno a medida que la sangre atraviesa los pulmones, luego la sangre atraviesa los capilares tisulares, su propia afinidad por el oxigeno permite que no lo libere los tejidos si ya hay demasiado; sin embargo si la concentración del oxigeno en el liquido tisular es demasiado baja, se libera oxigeno suficiente para restablecer una concentración adecuada.
La concentracion del dioxido de carbono (principal producto final metabólico de la celula) esta regulada de una forma diferente.
cuando hay demasiado dióxido en la sangre se estimula el centro respiratorio haciendo que la persona tenga una respiración rápida y profunda, este proceso continua hasta que la concentración vuelve a la normalidad.
Regulación de la presión arterial
varios sistemas nos ayudan a la regulacion de la presion arterial, por ejemplo los barroreceptores.
En las paredes de la zona en que se bifurcan las arterias carótidas en el cuello y también en el cayado torácico en el tórax, se encuentran los barroreceptores que se estimulan cuando se estira la pared arterial.
Cuando la presión arterial es elevada los barroreceptores se estimulan envían descargas de impulsos nerviosos al bulbo raquídeo cerebral y estos impulsos inhiben el centro vasomotor y disminuyen el numero de impulsos transmitidos desde el centro vasomotor a través del sistema nervioso simpático hacia el corazón y los vasos sanguíneos. La ausencia de estos impulsos hace que disminuya la actividad de bomba en el corazón y produce una dilatación de los vasos sanguíneos periféricos, lo que aumenta el flujo de sangre hasta que la presión arterial disminuya y recupere sus valores normales.
Si es caso es el contrario, el descenso de la presión arterial por debajo de lo normal relaja los receptores de estiramiento y hace que el centro vasomotor se vuelva mas activo de lo habitual, lo que provoca vasoconstriccion y aumento de la acción de la bomba cardíaca hasta alcanzar la normalidad.
Valores normales de los principales componentes en el liquido extracelular
En el presente trabajo se abordan el proceso general de curación de las heridas, la tipología y el tratamiento de las cicatrices y su tratamiento.
Los daños que llegan a sufrir los tejidos como consecuencia de agresiones físicas, químicas o biológicas, reciben el nombre genérico de lesiones.
Las lesiones presentan alteraciones más o menos profundas de la forma y función de los tejidos y órganos, que varían según el tipo de agente agresor, la magnitud del daño y la naturaleza del tejido o de los tejidos que resultaron afectados. Las heridas son lesiones ocasionadas por traumatismo mecánico en las que se observa rotura o interrupción de la continuidad de los tejidos blandos,3 y cuando el tejido lesionado es rígido o semirrígido, a la solución de la continuidad se le conoce como fractura. El conocimiento de la evolución de las heridas y de las fracturas es una de las bases teóricas más importantes en la educación médica,4 y dado que los mecanismos biológicos que hacen sanar las heridas son los mismos que conducen a la curación de las lesiones de origen químico o biológico en los diversos órganos y tejidos, el conocimiento del proceso es uno de los cimientos de la medicina contemporánea y objeto obligatorio de estudio en los primeros semestres de todas las carreras de las ciencias de la salud. A pesar de la gran cantidad de información acumulada y de los espectaculares avances en la biología celular y molecular, aún se sabe poco acerca de los mecanismos que regulan el proceso de la cicatrización, al grado de que con los recursos actuales, el cirujano se limita a no interferir y, en el mejor de los casos, a favorecer el proceso biológico que hace sanar las heridas sin que se conozca hasta el momento cómo regular su evolución. Para el cirujano, la herida que atiende llega a sanar en un proceso continuo y predecible, que finaliza cuando la lesión «cicatriza» y significa que se restableció la integridad física. En la investigación básica se considera el mismo proceso como una cascada armónica de hechos fisiológicos, a menudo difíciles de relacionar entre sí, que culmina con la creación de un nuevo órgano, la cicatriz, la cual evoluciona con el tiempo y continuará siendo objeto de estudio. En realidad no existe ninguna incongruencia en estos dos puntos de vista, puesto que cirujano y paciente observan efectos macroscópicos de un fenómeno biológico, en tanto que el investigador básico estudia los hechos que ocurren a nivel microscópico y molecular. El objetivo de este apartado es despertar el interés del estudiante para que su formación sea el resultado de la fusión de estos dos puntos de vista y que estén presentes en la esencia de todo aquel que se relacione con la cirugía, de modo que el cirujano tenga criterio de investigador para beneficio de los pacientes que atiende.
l. El proceso de sanar en los tejidos lesionados es una compleja cascada de eventos celulares, los cuales son coordinados por mediadores solubles y conducen a su restitución física y funcional.
La cicatrización en cada tejido tiene características propias, pero todos los tejidos sanan por mecanismos similares que cursan por las fases de inflamación, migración celular, proliferación, depósito de matriz y remodelación.
El cirujano debe conocer los factores locales, los factores sistémicos y las causas técnicas que pueden obstaculizar la evolución normal del proceso.
Los resultados óptimos se obtienen con la evaluación integral del paciente, de la herida y la aplicación de las mejores técnicas de práctica clínica.
La cicatrización anormal por exceso o por defecto plantea problemas clínicos en los que la genética, los factores del paciente y una buena técnica son determinantes.
Se espera que los avances en la comprensión de los factores de crecimiento, en la ingeniería de tejidos y de los materiales de curación enriquezcan el armamentario del cirujano.
Clasificación de las heridas según su causa Los medios mecánicos de agresión son múltiples, y entre ellos se encuentra el corte o incisión que hace el cirujano como parte del procedimiento. Las heridas se clasifican de acuerdo con la causa que las produce:
1. Heridas por instrumento punzocortante. Aquellas causadas por un objeto de borde filoso (como un cuchillo) o de extremidad aguda (como un clavo o punzón).
Heridas por contusión. Son ocasionadas cuando un objeto plano o de bordes redondeados golpea los tejidos blandos o cuando el cuerpo del individuo es proyectado con cierta velocidad sobre superficies planas que detienen de manera brusca su movimiento de aceleración.
Heridas por proyectil de arma de fuego. Los proyectiles acelerados por armas de fuego ocasionan lesiones complejas que difieren según las características del arma y de los propios proyectiles, los cuales pueden ser de alta velocidad y expansivos.
Heridas por machacamiento. Resultan cuando los tejidos son comprimidos entre dos superficies.
Heridas por laceración. Estas heridas se producen cuando los tejidos son arrancados.
Heridas por mordedura. Difieren en sus características y dependen de la especie animal que las produce. Entre las más comunes están las ocasionadas por otro humano, las cuales suelen inocularse con flora bacteriana múltiple; las mordeduras por cánidos suelen recibir cuidado especial por la posible transmisión del virus rábico. Las mordeduras por animales venenosos producen agresiones biológicas complejas.
Clasificación de las heridas según su profundidad
Excoriacion. Lesión superficial que afecta la epidermis y en general cicatriza regenerando en forma íntegra el epitelio, sin dejar huella visible.
Herida superficial. Es aquella que involucra a la piel y al tejido adiposo hasta la aponeurosis.
Herida profunda. Afecta los planos superficiales, la aponeurosis, (músculo y puede lesionar vasos, nervios y tendones.
Herida penetrante. Herida que lesiona los planos superficiales y llega al interior de las grandes cavidades. se les llama penetrante al abdomen, penetrante al wrax y penetrante al cráneo. En ocasiones hay dobles penetrantes.
Heridas según su estado bacteriológico
l. Herida 1ipo l. Herida limpia. Es la herida donde no hay contaminación exógena ni endógena, y en la que se supone que no habrá infección. Un ejemplo es la incisión que hace el cirujano
Herida tipo II. Herida limpia contaminada. Es una herida en la cual el cirujano sospecha que puede haber sufrido contaminación bacteriana, como donde hubo alguna violación de la técnica estéril del quirófano.
Herida tipo III. Herida contaminada. En este tipo de heridas se produjo una contaminación evidente, pero no están inflamadas ni tienen material purulento. Algunos ejemplos son las heridas como resultado de un traumatismo producido en la vía pública o en las intervenciones quirúrgicas donde ocurrió un derrame del contenido del tubo digestivo en la cavidad peritoneal, pero por ser recientes no tienen signos de infección activa.
Herida tipo IV. Herida :sucia o infectada. Es la herida que tiene franca infección evolutiva, por ejemplo, las que son resultado de un traumatismo más de 12 horas de haber sucedido, o la presencia de una fuente séptica muy bien identificada, como la perforación de una úlcera péptica o del apendice ileocecal con peritonitis purulenta, un absceso que se drena o un segmento de intestino gangrenado.
Hemostasia y fase inflamatoria Al producirse una herida hay un gran caos de células muertas, asi como sangre, cuerpos extraños y algunas bacterias. Para afrontar esta destrucción, la Naturaleza ha instrumentado una respuesta automática de defensa llamada inflamación.
TIPOS DE CICATRIZACION
–PRIMERA INTENCION: Todos los cirujanos que cierran una herida quisieran que cicatrizara por unión primaria o primera intención, con mínimo edema y sin infección local o secreción abundante. Una incisión que cicatriza por primera intención, lo hace en un tiempo mínimo, sin separación de los bordes de la herida, y con mínima formación de cicatriz. Esto se lleva a cabo en tres fases inflamación, proliferación, y remodelación.
–SEGUNDA INTENCION: Cuando la herida no cicatriza por unión primaria, se lleva a cabo un proceso de cicatrización más complicado y prolongado. La cicatrización por segunda intención es causada por infección, trauma excesivo, pérdida o aproximación imprecisa del tejido. En este caso, la herida puede dejarse abierta para permitir que cicatrice desde las capas profundas hacia la superficie exterior. Se forma tejido de granulación que contiene miofibroblastos y cierra por contracción. El proceso de cicatrización es lento y habitualmente se forma tejido de granulación y cicatriz. Como resultado, puede ser necesario que el cirujano trate el excesivo tejido de granulación que puede protruir por el margen de la herida y evitar epitelización.
–TERCERA INTENCION: También llamada cierre primario diferido, la cicatrización por tercera intención ocurre cuando dos superficies de tejido de granulación son aproximadas. Este es un método seguro de reparación de las heridas contaminadas, asó como de las heridas sucias e infectadas y traumatizadas, con pérdida extensa de tejido y riesgo elevado de infección. Este método se ha utilizado extensamente en el campo militar y ha probado que tiene éxito después de un trauma excesivo relacionado con accidentes automovilísticos, incidentes con armas de fuego, o heridas profundas y penetrantes con cuchillos. El cirujano habitualmente trata estas lesiones mediante debridación de los tejidos no viables y las deja abiertas. La herida abierta en cicatrización recupera gradualmente la suficiente resistencia a la infección que le permite un cierre no complicado. Generalmente esto se lleva a cabo cuatro a seis días después de la lesión. Este proceso se caracteriza por el desarrollo de yemas capilares y tejido de granulación. Cuando se lleva a cabo el cierre, los bordes de la piel y el tejido subyacente deben aproximarse y asegurarse con precisión.
–QUELOIDES: Las queloides también son conocidas como cicatrices con sobrecrecimiento. Se desarrollan un tiempo después de la cicatrización de la herida. El motivo es una producción excesiva de fibras de tejido conectivo, la cicatriz crece más allá de la herida y sobre la piel sana. Las cicatrices queloides acaban siendo más grandes que la herida original Las cicatrices en partes del cuerpo en las que la piel está sometida a una tensión elevada son susceptibles de formación de queloides. Con gran frecuencia, existe una predisposición genética a la formación de queloides. También se producen con una frecuencia diez veces superior en personas de piel más oscura.
–HIPERTROFICAS: Las cicatrices hipertróficas o elevadas se forman inmediatamente después de la cicatrización de la herida a causa de una producción excesiva de fibras de tejido conectivo. Este tipo de cicatriz tiende a sobresalir y destacar por encima del nivel de la piel circundante, pero permanece circunscrita a la región de la lesión original. Los factores de riesgo de las cicatrices elevadas son las infecciones y la inmovilización insuficiente de la herida.
–RETRACTIL O DEFORMANTE: Es la cicatriz fibrosa y extensa de los tejidos blandos que cubren las articulaciones o que están cerca de las articulaciones o que está cerca de los orificios naturales puede limitar los movimientos o incluso producir fijación permanente que ocasiona incapacidad física y consecuencias estéticas. Esta cicatriz distorsiona la forma y el tamaño de la piel es de importancia en la cara anterior del tórax y del cuello, la palma de la mano, las articulaciones en general, los parpados y los carrillos.
–CICATRICES ATROFICAS: Las cicatrices atróficas están ligeramente hundidas debido a la falta de colágeno. En estos casos, el tejido cicatrizal cubre la herida, pero no se produce suficiente tejido para cubrir toda la zona dañada. Las cicatrices atróficas son especialmente frecuentes después del acné o la varicela.
FASES DE CICATRIZACION
-FASE INFLAMATORIA: Para que se de esta fase primero al producirse la herida hay un gran desorden de células muertas, sangre, cuerpos extraños y algunas bacterias. Para preparar al cuerpo a este desorden se produce la inflamación que es la preparación del organismo que conduce a la curación de las heridas y una defensa contra otras lesiones o invasión, en esta etapa se ve ayudado por varios factores como las plaquetas que son fragmentos de células que intervienen en el proceso de coagulación, confluyen en mayor número al producirse una herida y liberan una serie de sustancias en la sangre, incluidas proteínas ECM, citoquinas, y factores de crecimiento. Los factores de crecimiento estimulan a las células para que aumenten su velocidad de división. Las plaquetas también liberan otros factores que favorecen la inflamación tales como son la serotonina, bradiquinina, prostaglandinas, prostaciclinas, tromboxano, e histamina; que aumentan la velocidad y migración de células hacia la zona, favorecen a los vasos sanguíneos en el proceso de dilatación y aumento de porosidad así como la activación de la cascada de coagulación que facilitara la cicatrización al promover la división celular.
FASE PROLIFERATIVA: Después de transcurridos dos a tres días desde la ocurrencia de la herida, comienza la afluencia de fibroblastos en la cicatriz, marcando el comienzo de la fase proliferativa aún antes que la fase inflamatoria haya concluido, por lo tanto en la proliferación reconstruye por lo que las fases no tienen una división cronológica y ocurren conjuntamente de manera armónica, la proliferación tiene elementos fundamentales:
1-epitelizacion 4-fibroplasia
2-angiogenesis 5-contraccion de la herida.
3-matriz de la herida
–FASE DE REMODELACION: Se realiza cuando ha sido reparada la rotura de la continuidad de los tejidos, el estimulo angiogeno disminuye en intensidad al parecer como una respuesta a las intenciones elevadas de oxígeno en los tejidos se inicia un periodo en el que la herida madura, presenta remodelación morfológica, reduce la hiperemia y su vascularidad y se reorganiza el tejido fibroso neoformado, a esto se le llama fase de remodelación y consiste en el descenso progresivo de los materiales formados en la cicatriz y en los cambios que experimenta en el tiempo.
Capítulo 78. Hormonas corticosuprarrenales
Las dos glándulas suprarrenales, con un peso aproximado de 4 g cada una, se hallan en los polos superiores de los riñones. cada glándula se compone de dos porciones diferentes, la médula suprarrenal y la corteza suprarrenal. La médula suprarrenal, que ocupa el 20% central de la glándula, se relaciona desde el punto de vista funcional con el sistema nervioso simpático; secreta las hormonas adrenalina y noradrenalina en respuesta a la estimulación simpática. A su vez, estas hormonas provocan casi los mismos efectos que la estimulación directa de los nervios simpáticos en todas las regiones del cuerpo.
Corticoesteroides: mineralocorticoides, glucocorticoides y andrógenos
La corteza suprarrenal secreta los dos tipos principales de hormonas corticosuprarrenales, los mineralocorticoides y los glucocorticoides. Además de estas hormonas, produce pequeñas cantidades de hormonas sexuales, en particular de andrógenos, que inducen los mismos efectos que la hormona sexual masculina testosterona. En general, son de escasa importancia, pero cuando se secretan en grandes proporciones en algunos trastornos de la corteza suprarrenal (como se expondrá más adelante este capítulo), causan los efectos virilizantes consiguientes. Los mineralocorticoides reciben este nombre porque afectan sobre todo a los electrólitos los minerales del compartimiento extracelular, especialmente al sodio y al potasio. Los glucocorticoides se denominan así porque poseen efectos importantes de aumento de la glucemia. Además, influyen en el metabolismo de las proteínas y de los lípidos, con efectos tan importantes para la función del organismo como los que producen sobre el metabolismo de los hidratos de carbono. Se han aislado más de 30 esteroides de la corteza suprarrenal, pero tan solo dos son determinantes para la función endocrina normal del cuerpo humano: la aldosterona, que es el mineralocorticoide principal, y el cortisol, que es el glucocorticoide principal.
La corteza suprarrenal tiene tres capas diferentes:
La zona glomerular, una capa delgada de células situada inmediatamente por debajo de la cápsula, contribuye con casi el 15% a la corteza suprarrenal. Estas células son las únicas de la glándula suprarrenal capaces de secretar cantidades importantes de aldosterona porque contienen la enzima aldosterona sintetasa, necesaria para la síntesis de la hormona. La secreción de estas células está controlada sobre todo por las concentraciones de angiotensina II y potasio en el líquido extracelular; ambos estimulan la secreción de aldosterona.
La zona fascicular, la zona media y más ancha, representa casi el 75% de la corteza suprarrenal y secreta los glucocorticoides cortisol y corticosterona, así como pequeñas cantidades de andrógenos y estrógenos suprarrenales. La secreción de estas células está controlada, en gran parte, por el eje hipotalámico-hipofisario a través de la corticotropina (ACTH).
La zona reticular, la capa más profunda de la corteza, secreta los andrógenos suprarrenales deshidroepiandrosterona (DHEA) y androstenodiona, así como pequeñas cantidades de estrógenos y algunos glucocorticoides. La ACTH también regula la secreción de estas células, aunque en ella pueden intervenir otros factores tales como la hormona corticótropa estimuladora de los andrógenos, liberada por la hipófisis. Sin embargo, los mecanismos que regulan la producción suprarrenal de andrógenos no se conocen tan bien como los de los glucocorticoides y mineralocorticoides.
Todas las hormonas esteroideas humanas, incluidas las producidas por la corteza suprarrenal, se sintetizan a partir del colesterol, Si bien las células de la corteza suprarrenal pueden sintetizar de novo pequeñas cantidades de colesterol a partir del acetato, casi el 80% del colesterol empleado para la síntesis de esteroides proviene de las lipoproteínas de baja densidad (LDL) del plasma circulante. Las LDL, que transportan altas concentraciones de colesterol, difunden desde el plasma al líquido intersticial para unirse a receptores específicos localizados en estructuras de la membrana de la célula corticosuprarrenal conocidas como depresiones revestidas. Estas depresiones penetran en el citoplasma por endocitosis, transformándose en vesículas que, por último, se fusionan con los lisosomas y liberan el colesterol destinado a la síntesis de los esteroides suprarrenales. El transporte del colesterol a las células suprarrenales está sometido a mecanismos de retroalimentación que pueden modificar en gran medida la cantidad disponible para la síntesis de esteroides. Por ejemplo, la ACTH, que estimula la síntesis de esteroides suprarrenales, incrementa el número de receptores de LDL de la célula corticosuprarrenal y la actividad de las enzimas que liberan el colesterol a partir de las LDL.
Etapas principales de la síntesis de los productos esteroideos más importantes de la corteza suprarrenal: aldosterona, cortisol y andrógenos.
Casi todas estas etapas suceden en dos orgánulos celulares, las mitocondrias y el retículo endoplásmico, pero algunas tienen lugar en las primeras y otras en el segundo. Cada etapa está catalizada por un sistema enzimático específico. Un cambio, incluso de una sola enzima, puede provocar la formación de tipos muy distintos y porcentajes diferentes de hormonas. Por ejemplo, si se altera la actividad de tan solo una enzima de esta vía, se generarán cantidades enormes de hormonas sexuales masculinizantes u otros compuestos esteroideos que normalmente no se encuentran en la sangre. Las fórmulas químicas de la aldosterona y el cortisol, que son las principales hormonas mineralocorticoide y glucocorticoide, respectivamente. la corteza suprarrenal suele secretar pequeñas cantidades de otros esteroides con actividad glucocorticoide, mineralocorticoide o mixta. Por último, se han sintetizado y empleado en diversas formas de tratamiento varias hormonas esteroideas potentes, no sintetizadas por las glándulas suprarrenales en condiciones normales.
Mineralocorticoides
Aldosterona (muy potente, supone casi el 90% de toda la actividad mineralocorticoide).
Desoxicorticosterona (1/30 de la potencia de la aldosterona, aunque se secreta en cantidades mínimas).
9α-fluorocortisol (sintético, algo más potente que la aldosterona).
Cortisol (actividad mineralocorticoide mínima, pero se secreta en grandes cantidades).
Cortisona (actividad mineralocorticoide mínima). Glucocorticoides • Cortisol (muy potente; es el responsable de casi el 95% de toda la actividad glucocorticoide).
Corticosterona (proporciona el 4% de la actividad glucocorticoide total, pero es mucho menos potente que el cortisol).
Cortisona (casi tan potente como el cortisol).
Prednisona (sintética, cuatro veces más potente que el cortisol).
Metilprednisolona (sintética, cinco veces más potente que el cortisol).
Dexametasona (sintética, 30 veces más potente que el cortisol).
Funciones de los mineralocorticoides: aldosterona La deficiencia de mineralocorticoides provoca pérdidas renales intensas de cloruro sódico e hiperpotasemia
La pérdida completa de la secreción corticosuprarrenal puede causar la muerte en un plazo de 3 días a 2 semanas, salvo que la persona reciba un tratamiento salino intensivo o la inyección de mineralocorticoides. Sin mineralocorticoides, la concentración del ion potasio del líquido extracelular experimenta un gran ascenso, el sodio y el cloruro desaparecen enseguida del organismo y el volumen total del líquido extracelular y el volumen de sangre se reducen mucho. Pronto se desarrolla un descenso del gasto cardíaco, que evoluciona a un estado de shock, seguido de la muerte. Toda esta secuencia puede evitarse con la administración de aldosterona u otro mineralocorticoide. En el ser humano, la aldosterona es la responsable de casi el 90% de la actividad mineralocorticoide de las secreciones corticosuprarrenales, pero el cortisol, el principal glucocorticoide secretado por la corteza suprarrenal, también aporta una actividad mineralocorticoide importante. La actividad mineralocorticoide de la aldosterona es alrededor de 3.000 veces mayor que la del cortisol, pero la concentración plasmática de este último es casi 2.000 veces superior a la de la aldosterona. El cortisol puede unirse asimismo a receptores mineralocorticoides con alta afinidad. Sin embargo, las células epiteliales renales expresan la enzima 11β-hidroxiesteroide deshidrogenasa de tipo 2 (11βHSD2), que tiene acciones que evitan que el cortisol active los receptores mineralocorticoides. Una acción de la 11β-HSD2 consiste en convertir el cortisol en cortisona, que no muestra avidez por unirse a receptores mineralocorticoides.
La aldosterona ejerce casi los mismos efectos sobre las glándulas sudoríparas y salivales que sobre los túbulos renales.
Estos dos tipos de glándulas producen una secreción primaria que contiene grandes cantidades de cloruro sódico, aunque gran parte del cloruro sódico se reabsorbe al atravesar los conductos excretores, mientras que los iones potasio y bicarbonato se excretan. La aldosterona aumenta de manera considerable la reabsorción de cloruro sódico y la secreción de potasio por los conductos. El efecto sobre las glándulas sudoríparas reviste interés para conservar la sal del organismo en ambientes cálidos y el efecto sobre las glándulas salivales permite conservar la sal cuando se pierden cantidades excesivas de saliva. Desde hace muchos años se conocen los efectos generales de los mineralocorticoides sobre el organismo, pero se ignora el mecanismo molecular de la acción de la aldosterona incrementa el transporte de sodio en las células tubulares. No obstante, la secuencia celular de acontecimientos que culmina con el aumento de la reabsorción de sodio parece ser la siguiente.
En primer lugar, la aldosterona difunde de inmediato al interior de las células del epitelio tubular, debido a su liposolubilidad en las membranas celulares.
En segundo lugar, la aldosterona se une a la proteína receptor mineralocorticoide (MR), una proteína que dispone de una configuración estereomolecular por la que solo la aldosterona o compuestos muy parecidos se unen a ella. Aunque los receptores MR de células epiteliales tubulares renales también poseen una alta afinidad por el cortisol, la enzima 11β-HSD2 convierte normalmente la mayor parte del cortisol en cortisona, que no se une fácilmente a los receptores MR, como se expone anteriormente.
En tercer lugar, el complejo aldosterona-receptor o algún producto de este complejo difunde al interior del núcleo, donde sufre nuevas alteraciones para, por último, inducir la síntesis de uno o más tipos de ARN mensajero (ARNm) relacionados con el transporte del sodio y del potasio. En cuarto lugar, el ARNm pasa al citoplasma, donde, en colaboración con los ribosomas, induce la formación de proteínas.
Las proteínas así formadas consisten en:
una o más enzimas
proteínas transportadoras de membrana, cuya presencia conjunta es imprescindible para el transporte de sodio, potasio e hidrógeno a través de la membrana celular.
La regulación de la secreción de aldosterona está tan íntimamente ligada al control de las concentraciones de electrólitos en el líquido extracelular, el volumen del líquido extracelular, el volumen sanguíneo, la presión arterial y muchos aspectos especiales de la función renal que resulta difícil exponerla con independencia de todos ellos. Este tema se expuso con más detalle en los capítulos 29 y 30, a los que se remite al lector. Sin embargo, conviene enumerar aquí los aspectos más relevantes del control de la secreción de aldosterona. La regulación de la secreción de aldosterona por las células de la zona glomerular no depende apenas de la regulación del cortisol o de los andrógenos por las zonas fascicular y reticular.
Se conocen los siguientes cuatro factores que desempeñan una función esencial para la regulación de la aldosterona:
El incremento de la concentración de iones potasio en el líquido extracelular aumenta mucho la secreción de aldosterona.
El aumento de la concentración de angiotensina II en el líquido extracelular también incrementa mucho la secreción de aldosterona.
El incremento de la concentración de iones sodio en el líquido extracelular apenas reduce la secreción de aldosterona.
Se necesita ACTH de la adenohipófisis para que haya secreción de aldosterona, aunque su efecto regulador sobre la velocidad de secreción es mínimo en la mayoría de los trastornos fisiológicos.
La glándula tiroides, situada justo por debajo de la laringe y a ambos lados y por delante de la tráquea, es una de las glándulas endocrinas más grandes, con un peso que oscila entre 15 y 20 g en los adultos sanos. El tiroides secreta dos hormonas importantes, la tiroxina y la triyodotironina, conocidas a menudo como T4 y T3, respectivamente. Ambas inducen un notable aumento del metabolismo del organismo. La ausencia completa de secreción tiroidea provoca con frecuencia descensos metabólicos de hasta un 40-50% inferiores al valor normal, mientras que la secreción excesiva incrementa el metabolismo en hasta el 60-100% por encima de lo normal. La secreción tiroidea está controlada por la tirotropina (TSH), secretada por la adenohipófisis.
Síntesis y secreción de las hormonas metabólicas tiroideas
Alrededor del 93% de las hormonas con actividad metabólica secretadas por la glándula tiroides corresponde a tiroxina y el 7% restante, a triyodotironina. No obstante, con el tiempo, casi toda la tiroxina se convierte en triyodotironina en los tejidos, por lo que ambas desempeñan funciones importantes. Estas funciones son cualitativamente similares, aunque difieren en la rapidez y la intensidad de la acción. La triyodotironina es unas cuatro veces más potente que la tiroxina, si bien se detecta una cantidad mucho menor en la sangre y su duración es más breve.
Anatomía fisiológica de la glándula tiroides
La glándula tiroides se compone de un elevado número de folículos cerrados (100 a 300 μm de diámetro), que están repletos de una sustancia secretora denominada coloide y revestidos por células epiteliales cúbicas que secretan a la luz de los folículos. El componente principal del coloide es una glucoproteína de gran tamaño, la tiroglobulina, cuya molécula contiene las hormonas tiroideas. Cuando la secreción se encuentra en los folículos, la sangre debe absorberla de nuevo a través del epitelio folicular para que pueda actuar en el organismo.
Para formar una cantidad normal de tiroxina se precisan al año unos 50 mg de yodo (ingerido en forma de yoduros) o el equivalente a 1 mg/semana. Para impedir la deficiencia de yodo, se añade una parte de yoduro sódico por cada 100.000 partes de cloruro sódico a la sal de mesa común. Los yoduros ingeridos por vía oral se absorben desde el tubo digestivo hasta la sangre de la misma forma que los cloruros. En condiciones normales, la mayor parte se excreta con rapidez por vía renal, pero siempre después de que las células tiroideas hayan retirado selectivamente una quinta parte de la sangre circulante y la hayan empleado en la síntesis de las hormonas tiroideas.
Bomba de yoduro: el simportador del yoduro de sodio (atrapamiento de yoduro) La primera etapa de la formación de las hormonas tiroideas consiste en el transporte de los yoduros desde la sangre hasta las células y los folículos de la glándula tiroides. La membrana basal de estas células posee la capacidad específica de bombear de forma activa el yoduro al interior celular. Este bombeo se consigue mediante la acción de un simportador del yoduro de sodio, que cotransporta el ion yoduro a lo largo de dos iones sodio a través de la membrana basolateral (plasma) a la célula. La energía para el transporte del yoduro en contra de un gradiente de concentración proviene de la bomba de sodio-potasio-adenosina trifosfatasa (ATPasa), que bombea sodio al exterior de la célula, con lo que establece una baja concentración de sodio intracelular y un gradiente para facilitar la difusión de sodio en la célula.
En una glándula normal, la bomba de yoduro concentra esta sustancia hasta que su concentración supera en 30 veces la de la sangre. Cuando la glándula tiroides alcanza su máxima actividad, la relación entre ambas concentraciones puede elevarse hasta 250 veces. El atrapamiento de yoduro por la glándula tiroides depende de diversos factores, el más importante de los cuales es la concentración de TSH; esta hormona estimula la actividad de la bomba de yoduro en las células tiroideas, mientras que la hipofisectomía la disminuye. El yoduro es transportado fuera de las células tiroideas a través de la membrana apical hacia el folículo por una molécula de contratransporte de cloruro-yoduro denominada pendrina.
Tiroglobulina y química de la formación de tiroxina y triyodotironina.
Formación y secreción de tiroglobulina por las células tiroideas Las células tiroideas constituyen un ejemplo típico de células glandulares secretoras de proteínas. El retículo endoplásmico y el aparato de Golgi sintetizan y secretan hacia los folículos una gran molécula glucoproteica denominada tiroglobulina, con un peso molecular aproximado de 335.000. Cada molécula de tiroglobulina contiene unas 70 moléculas del aminoácido tirosina, que es el sustrato principal que se combina con el yodo para dar lugar a las hormonas tiroideas. Así pues, las hormonas tiroideas se forman dentro de la molécula de tiroglobulina.
El primer paso crítico para la formación de las hormonas tiroideas consiste en la conversión de los iones yoduro en una forma oxidada del yodo, bien en yodo naciente (I 0 ), bien en I 3 – , que luego puede combinarse directamente con el aminoácido tirosina. La oxidación del yodo depende de la enzima peroxidasa y su peróxido de hidrógeno acompañante, que constituyen un potente sistema capaz de oxidar los yoduros. La peroxidasa se encuentra en la membrana apical de la célula o unida a ella, proporcionando así el yodo oxidado justo en el lugar de la célula donde la molécula de tiroglobulina abandona el aparato de Golgi y atraviesa la membrana celular hasta el coloide almacenado en la glándula tiroides. Cuando el sistema de la peroxidasa se bloquea o en los casos de ausencia congénita, la velocidad de formación de hormonas tiroideas disminuye hasta cero.
Yodación de la tirosina y formación de las hormonas tiroideas: «organificación» de la tiroglobulina La unión del yodo a la molécula de tiroglobulina recibe el nombre de organificación de la tiroglobulina. El yodo oxidado (incluso en forma molecular) se une directamente, aunque con lentitud, al aminoácido tirosina. No obstante, en las células tiroideas el yodo oxidado se asocia a la enzima tiroidea peroxidasa, que hace que el proceso tenga lugar en segundos o minutos. Por consiguiente, a medida que la tiroglobulina se libera del aparato de Golgi o se secreta al folículo a través de la membrana apical de la célula, el yodo se fija a alrededor de la sexta parte de las tirosinas contenidas en la molécula de tiroglobulina.
El principal producto hormonal de la reacción de acoplamiento es la molécula tiroxina (T4), que se forma cuando se unen dos moléculas de diyodotirosina; la tirosina forma parte aún de la molécula de tiroglobulina. En otras ocasiones, una molécula de monoyodotirosina se une con una de diyodotirosina para formar triyodotironina (T3), que representa alrededor de la quinceava parte del total final de hormonas. Se forman pequeñas cantidades de T3 inversa (RT3) mediante acoplamiento de diyodotirosina con monoyodotirosina, aunque la RT3 no parece tener importancia funcional en los seres humanos. Almacenamiento de la tiroglobulina La glándula tiroides es la única glándula endocrina que posee la capacidad de almacenar grandes cantidades de hormona. Una vez finalizada la síntesis de las hormonas tiroideas, cada molécula de tiroglobulina contiene hasta 30 moléculas de tiroxina y algunas de triyodotironina.
Liberación de tiroxina y triyodotironina del tiroides
La mayor parte de la tiroglobulina no se libera a la sangre circulante, sino que es preciso que la tiroxina y triyodotironina se escindan de la molécula de tiroglobulina; a continuación, ambas se secretan en forma libre. Este proceso tiene lugar por el siguiente mecanismo: la superficie apical de las células tiroideas emite extensiones en forma de seudópodos que rodean a pequeñas porciones del coloide, constituyendo vesículas de pinocitosis, que alcanzan la punta de la célula tiroidea. A continuación, los lisosomas del citoplasma celular se funden de inmediato con estas vesículas y forman otras vesículas digestivas que contienen enzimas procedentes de los lisosomas mezcladas con el coloide. Varias enzimas proteinasas digieren las moléculas de tiroglobulina, y liberan la tiroxina y la triyodotironina, que se difunden entonces a través de la base de la célula tiroidea, hacia los capilares circundantes, y de este modo pasan a la sangre. Parte de la tiroglobulina del coloide entra en la célula tiroidea por endocitosis después de su unión a la megalina, una proteína situada en la membrana luminal de las células. A continuación, el complejo megalina-tiroglobulina es transportado a través de la célula por transcitosis hasta la membrana basolateral, donde una parte de la megalina permanece unida a la tiroglobulina y es liberada en la sangre capilar. Alrededor de las tres cuartas partes de la tirosina yodada en la tiroglobulina nunca se convierten en hormona tiroidea, sino que permanecen como monoyodotirosina y diyodotirosina.
Transporte de tiroxina y triyodotironina a los tejidos La tiroxina y la triyodotironina están unidas a proteínas plasmáticas Cuando acceden a la sangre, más del 99% de la tiroxina y la triyodotironina se combina de inmediato con diversas proteínas plasmáticas, todas ellas sintetizadas por el hígado. Estas proteínas son, ante todo, la globulina fijadora de la tiroxina y, en menor medida, la prealbúmina y la albúmina fijadora de la tiroxina. La tiroxina y la triyodotironina se liberan lentamente a las células de los tejidos Debido a la gran afinidad de las proteínas de unión plasmáticas por las hormonas tiroideas, estas sustancias, en concreto la tiroxina, se liberan con lentitud a las células de los tejidos. La mitad de la tiroxina presente en la sangre se libera a las células de los tejidos cada 6 días aproximadamente, mientras que la mitad de la triyodotironina, dada su menor afinidad, tarda 1 día en llegar a las células.
La glándula suprarrenal consta de dos porciones diferentes: 1) una médula interna, relacionada funcionalmente con el sistema nervioso simpático, que segrega principalmente adrenalina, pero también algo de noradrenalina, y 2) una corteza externa, que constituye la mayor parte de la glándula y segrega corticoesteroides. Los principales corticoesteroides de la corteza suprarrenal son los siguientes: Mineralocorticoides. Esteroides C21 con importantes efectos sobre el balance de sodio y potasio.
Glucocorticoides. Esteroides C21 que modifican el metabolismo de los hidratos de carbono, grasas y proteínas. Hormonas sexuales. Esteroides C19, en su mayoría andrógenos débiles, que contribuyen a las características sexuales secundarias. La secreción de mineralocorticoides y glucocorticoides resulta esencial para la vida. La corteza suprarrenal segrega normalmente pequeñas cantidades de hormonas sexuales, con un efecto mínimo sobre la función reproductora.
QUÍMICA DE LA SECRECIÓN CORTICOSUPRARRENAL
La corteza suprarrenal se compone de tres capas o tipos de células diferentes: zona glomerular, zona fascicular y zona reticular: La zona glomerular, o externa, es bastante delgada y representa el lugar exclusivo de la enzima aldosterona sintasa. El producto de secreción principal es el mineralocorticoide más importante, la aldosterona. La angiotensina JI y el potasio son los principales reguladores de la secreción de aldosterona. Las elevaciones crónicas de la concentración plasmática de la angiotensina II, que ocurren, por ejemplo, cuando disminuye el sodio, producen hipertrofia e hiperplasia, pero solo de las células de la zona glomerular.
Dado que esta zona carece de la enzima 17-hidroxilasa , no puede sintetizar cortisol ni hormonas sexuales. La zona fascicular, o intermedia, es la más ancha; segrega los glucocorticoides cortisol (el glucocorticoide principal) y corticosterona. Esta zona también segrega pequeñas cantidades de hormonas sexuales. La hormona adrenocorticótropa (corticotropina, ACTH) es la reguladora más importante de la secreción de cortisol. La zona reticular, o interna, segrega las hormonas sexuales y algunos glucocorticoides; como la zona fascicular, es estimulada por la ACTH. El exceso crónico de ACTH produce hipertrofia e hiperplasia de las dos zonas internas de la corteza suprarrenal. Los andrógenos suprarrenales más prevalentes son la deshidroepiandrosterona (DHEA) y la androstenodiona.
Las hormonas corticosuprarrenales se sintetizan a partir del colesterol. Casi todo el colesterol de las células corticosuprarrenales es captado de la circulación y posteriormente esterificado y almacenado en gotitas lipídicas. El paso que limita la velocidad de síntesis de las hormonas corticosuprarrenales es la escisión de la cadena lateral del colesterol para formar pregnenolona.
Este paso incluye la liberación de colesterol a la membrana mitocondrial interna y el desdoblamiento enzimático (por la colesterol desmolasa) de una unidad de seis carbonos de colesterol para obtener pregnenolona. Los reguladores de los principales productos hormonales (aldosterona y cortisol) estimulan este paso inicial en la biosíntesis de los esteroides en las tres zonas de la corteza suprarrenal. La conversión del colesterol en pregnenolona y todas las etapas sucesivas de la síntesis de las hormonas corticosuprarrenales tienen lugar en el retículo endoplásmico o en las mitocondrias. No todos los compuestos se producen en las tres zonas de las cortezas suprarrenal.
Las hormonas corticosuprarrenales están unidas a las proteínas plasmáticas. Del 90 al 95% del cortisol plasmático se une a las proteínas del plasma, principalmente a la transcortina o globulina fijadora de los corticoesteroides. El cortisol posee una semivida prolongada (de 60 a 90 min) como consecuencia de su elevada unión a las proteínas del plasma. La corticosterona se une menos a ellas que el colesterol, y su semivida se aproxima a 50 min. La aldosterona se liga todavía menos a las proteínas del plasma y, en consecuencia, su semivida solo llega a 20 min. Las hormonas corticosuprarrenales se metabolizan en el hígado. El cortisol y la aldosterona se metabolizan hacia diversos compuestos en el hígado y luego se conjugan con el ácido glucurónico. Estos conjugados inactivos son fácilmente solubles en el plasma y no se unen a sus proteínas. Una vez liberados a la circulación, se excretan con facilidad a través de la orina. La velocidad de inactivación de las hormonas corticosuprarrenales disminuye en las hepatopatías.
FUNCIONES DE LOS MINERALOCORTICOIDES: ALDOSTERONA
La aldosterona es el mineralocorticoide principal segregado por la corteza suprarrenal. La aldosterona da cuenta de casi el 90% de la actividad mineralocorticoide de las hormonas corticosuprarrenales. Casi todo el resto de la actividad mineralocorticoide se debe a: 1) la desoxicorticosterona, que posee un 3% de la actividad rnineralocorticoide de la aldosterona y se segrega con un ritmo similar, y 2) el cortisol, un grupo corticoide con una débil actividad mineralocorticoide, presente en condiciones normales en el plasma en concentraciones más de 1.000 veces mayores que la aldosterona.
La aldosterona también hace que se segreguen iones hidrógeno, que se intercambian por sodio, en las células intercaladas de los túbulos colectores corticales. Para que ocurran las acciones tubulares de la aldosterona se requiere la síntesis de proteínas, lo que explica la latencia de unos 60 min entre la exposición a la aldosterona y el comienzo de sus efectos. La aldosterona modifica el transporte de los electrólitos en órganos distintos de los riñones. La aldosterona se une a los receptores mineralocorticoides de las células epiteliales, diferentes a los renales. La aldosterona aumenta la reabsorción de sodio en el colon y fomenta la excreción de potasio con las heces. De forma análoga, la aldosterona posee un efecto sobre las glándulas sudoríparas y salivales, puesto que reduce el cociente sodio/potasio en las secreciones respectivas.
Regulación de la secreción de aldosterona por la angiotensina II y el potasio
La angiotensina 11 estimula la secreción de aldosterona. La angiotensina JI estimula directamente las células de la zona glomerular para que segreguen aldosterona. Este efecto de la angiotensina ll está mediado a través de incrementos en los niveles intracelulares de calcio y los productos del fosfatidilinositol diacilglicerol y trifosfato de inositol. Estos segundos mensajeros activan la proteína cinasa C, que, a su vez, estimula los pasos temprano (colesterol desmolasa) y tardío (aldosterona sintasa) para la biosíntesis de la aldosterona. El control de la secreción de aldosterona por la angiotensina ll está íntimamente asociado a la regulación del volumen extracelular y de la presión arterial. El sistema renina-angiotensina se activa en presencia de hipovolemia e hipotensión; los valores plasmáticos elevados de angiotensina II estimulan la secreción de aldosterona. A su vez, la aldosterona aumenta la reabsorción de sodio en la porción distal de la nefrona; en cuanto la retención de líquidos restaura el volumen de los líquidos corporales y la presión arterial a la normalidad, desaparece el estímulo para la activación del sistema renina-angiotensina y la secreción de aldosterona desciende hasta los valores basales.
La aldosterona desempeña una misión fundamental para eliminar el potasio ingerido y regular, mediante retroalimentación, la concentración plasmática de potasio. Los aumentos en la concentración plasmática de potasio elevan la secreción de aldosterona, que, a su vez, estimula la secreción tubular de potasio. Conforme la concentración plasmática de potasio disminuye hasta cifras normales, desaparece el estímulo para la secreción de aldosterona. Cuando la concentración plasmática de potasio disminuye, ocurre una secuencia antagónica de acontecimientos. El incremento en la concentración plasmática de potasio despolariza la membrana celular, activando los canales de calcio dependientes del voltaje. La elevación del calcio citoplásmico estimula la secreción de aldosterona por el mecanismo descrito más arriba para la angiotensina 11.
La ACTH ejerce una función permisiva en la regulación de la secreción de aldosterona. Mientras las cifras plasmáticas de ACTH se mantengan normales, la capacidad de respuesta de la zona glomerular a los principales elementos reguladores, la angiotensina II y el potasio, se preserva. En cambio, cuando existe una deficiencia crónica de ACTH, disminuye la respuesta de la aldosterona a la angiotensina II y al potasio. Los niveles plasmáticos elevados de ACTH, que se acentúan de manera aguda durante el estrés, estimulan la secreción de aldosterona; no obstante, en los estados de exceso crónico de ACTH (p. ej., enfermedad de Cushing), no se perpetúa el hiperaldosteronismo.
Regulación de la secreción de cortisol por la ACTH
La ACTH estimula la secreción de cortisol. La secreción de cortisol está sujeta a la regulación del eje hipotálamohipófisis: hormona liberadora de corticotropina (CRH)- ACTH. La liberación de ACTH (corticotropina) por la hipófisis depende de la hormona hipofisótropa CRH. En cuanto se segrega ACTH a la sangre, actúa rápidamente sobre las dos zonas internas de la corteza suprarrenal, principalmente sobre la zona fascicular, lo que produce un incremento de la secreción de cortisol.
El estrés aumenta la secreción de ACTH. Diversos factores físicos y mentales de estrés estimulan las células neuroendocrinas del hipotálamo para que segreguen CRH; el resultado es un aumento en la secreción de la ACTH, que estimula la liberación de cortisol. En los estados de estrés, el efecto inhibitorio del cortisol sobre la secreción de ACTH no basta para contrarrestar las aferencias neurales adicionales que alcanzan las células neuroendocrinas y fomentan la secreción de CRH. Por tanto, los valores plasmáticos de la ACTH se elevan.
ANOMALÍAS DE LA SECRECIÓN CORTICOSUPRARRENAL
El aumento en los niveles plasmáticos de los glucocorticoides (cortisol) produce el síndrome de Cushing. La secreción excesiva de cortisol puede ser producida por un tumor suprarrenal, un tumor hipofisario que segregue grandes cantidades de ACTH y produzca una hiperplasia suprarrenal bilateral (síndrome de Cushing), o un tumor de los pulmones,u otros tejidos (un tumor ectápico) que segreguen grandes cantidades de ACTH y produzcan hiperplasia bilateral. El síndrome de Cushing también puede deberse a la administración de grandes cantidades de glucocorticoides por vía exógena.
El síndrome de Conn (aldosteronismo primario) es producido por un tumor de la zona glomerular. Si hay un tumor en esta zona, que produce grandes cantidades de aldosterona, las manifestaciones más características son la hipertensión y la hipopotasemia. En general, la hipertensión es bastante leve, porque solo ocurre un pequeño aumento del volumen extracelular debido al «escape de sodio». La hipertensión y la hipopotasemia se exacerban con el aumento del aporte de sodio. Dada la expansión en el volumen extracelular y el aumento de la presión arterial, la actividad plasmática de la renina se suprime. El descenso del potasio en el síndrome de Conn reduce la capacidad de concentración de los riñones, produce poliuria, y ocasiona debilidad muscular y alcalosis metabólica.
la corriente eléctrica también se propaga desde el corazón hacia los tejidos adyacentes que lo rodean. Una pequeña parte de la corriente se propaga hacia la superficie corporal. Si se colocan electrodos en la piel en lados opuestos del corazón se pueden registrar los potenciales eléctricos que se generan por la corriente; el registro se conoce como electrocardiograma (ECG)
Características del electrocardiograma normal El ECG normal (v. fig. 11-1) está formado por una onda P, un complejo QRS y una onda T. Con frecuencia, aunque no siempre, el complejo QRS está formado por tres ondas separadas: la onda Q, la onda R y la onda S. La onda P está producida por los potenciales eléctricos que se generan cuando se despolarizan las aurículas antes del comienzo de la contracción auricular. El complejo QRS está formado por los potenciales que se generan cuando se despolarizan los ventrículos antes de su contracción, es decir, a medida que la onda de despolarización se propaga por los ventrículos. Por tanto, tanto la onda P como los componentes del complejo QRS son las ondas de despolarización. La onda T está producida por los potenciales que se generan cuando los ventrículos se recuperan del estado de despolarización. Este proceso normalmente aparece en el músculo ventricular entre 0,25 y 0,35 s después de la despolarización. La onda T se conoce como onda de repolarización. Así, el ECG está formado por ondas tanto de despolarización como de repolarización. La distinción entre ondas de despolarización y ondas de repolarización es tan importante en electrocardiografía que requiere una aclaración adicional.
Ondas de despolarización frente a ondas de repolarización
Durante la despolarización el potencial negativo normal del interior de la fibra se invierte y se hace ligeramente positivo en el interior y negativo en el exterior.
En la figura A, la despolarización, que se indica por las cargas positivas de color rojo del interior y las cargas negativas de color rojo del exterior, se dirige desde la izquierda hacia la derecha. La primera mitad de la fibra ya se ha despolarizado, mientras que la mitad restante sigue polarizada. Por tanto, el electrodo izquierdo del exterior de la fibra está en una zona de negatividad, y el electrodo derecho está en una zona de positividad, lo que hace que el medidor registre un valor positivo. A la derecha de la fibra muscular se muestra un registro de los cambios de potencial entre los dos electrodos, que se registran con un medidor de registro de alta velocidad. cuando la despolarización ha alcanzado la marca intermedia el registro ha aumentado hasta un valor positivo máximo. En la figura B la despolarización se ha propagado por toda la fibra muscular, y el registro de la derecha ha vuelto a la línea basal de cero porque los dos electrodos ahora están en zonas de igual negatividad. La onda completa es una onda de despolarización porque se debe a la propagación de la despolarización a lo largo de la membrana de la fibra muscular. La figura C muestra la mitad de la repolarización de la misma fibra muscular, de modo que vuelve la positividad al exterior de la fibra. En este punto el electrodo izquierdo está en una zona de positividad y el electrodo derecho está en una zona de negatividad. Esta polaridad es opuesta a la polaridad de la figura A. Por tanto, el registro, que se muestra a la derecha, se hace negativo. En la figura D la fibra muscular se ha repolarizado completamente, y los dos electrodos están ahora en zonas de positividad, de modo que no se registra ninguna diferencia de potencial entre ellos. Por tanto, en el registro de la derecha el potencial vuelve una vez más a cero. Esta onda negativa completa es una onda de repolarización porque se debe a la propagación de la repolarización a lo largo de la membrana de la fibra muscular.
Relación del potencial de acción monofásico del músculo ventricular con las ondas QRS y T del electrocardiograma estándar
un potencial de acción monofásico registrado con un microelectrodo insertado en el interior de una fibra muscular ventricular única. El ascenso de este potencial de acción está producido por la despolarización, y la vuelta del potencial al nivel basal está producida por la repolarización
Relación de la contracción auricular y ventricular con las ondas del electrocardiograma
Antes de que se pueda producir la contracción del músculo, la despolarización se debe propagar por todo el músculo para iniciar los procesos químicos de la contracción. La onda P se produce al comienzo de la contracción de las aurículas y el complejo QRS de ondas se produce al comienzo de la contracción de los ventrículos. Los ventrículos siguen contraídos hasta después de que se haya producido la repolarización, es decir, hasta después del final de la onda T. Las aurículas se repolarizan aproximadamente 0,15 a 0,2 s después de la finalización de la onda P, lo que coincide aproximadamente con el momento en el que se registra el complejo QRS en el ECG. Por tanto, la onda de repolarización auricular, conocida como onda T auricular, habitualmente está oscurecida por el complejo QRS, que es mucho mayor. Por este motivo raras veces se observa la onda T auricular en el ECG. La onda de repolarización ventricular es la onda T del ECG normal. Habitualmente el músculo ventricular comienza a repolarizarse en algunas fibras aproximadamente 0,2 s después del comienzo de la onda de despolarización (el complejo QRS), pero en muchas otras fibras tarda hasta 0,35 s. Así, el proceso de repolarización ventricular se extiende a lo largo de un período prolongado, de aproximadamente 0,15 s. Por este motivo la onda T del ECG normal es una onda prolongada, aunque el voltaje de la onda T es mucho menor que el voltaje del complejo QRS, en parte debido a esta duración prolongada.
Calibración del voltaje y el tiempo del electrocardiograma Todos los registros de los ECG se hacen con líneas de calibración adecuadas sobre el papel de registro. Estas líneas de calibración pueden estar ya señaladas en el papel, como ocurre cuando se utiliza un registrador de pluma, o se registran en el papel al mismo tiempo que se registra el ECG, como en los tipos fotográficos de electrocardiógrafos.
Voltajes normales en el electrocardiograma Los voltajes de las ondas que se registran en el ECG normal dependen de la manera en la que se aplican los electrodos a la superficie del cuerpo y de la proximidad de los electrodos al corazón. Cuando un electrodo está colocado directamente sobre los ventrículos y un segundo electrodo está localizado en otra localización del cuerpo alejada del corazón, el voltaje del complejo QRS puede ser de hasta 3 a 4 mV. Incluso este voltaje es pequeño en comparación con el potencial de acción monofásico de 110 mV que se registra directamente en la membrana del músculo cardíaco. Cuando los ECG se registran con electrodos en los dos brazos o en un brazo y una pierna, el voltaje en el complejo QRS habitualmente es de 1 a 1,5 mV desde el punto más elevado de la onda R hasta el punto más profundo de la onda S; el voltaje de la onda P está entre 0,1 y 0,3 mV, y el de la onda T está entre 0,2 y 0,3 mV.
Intervalo P-Q o P-R El tiempo que transcurre entre el comienzo de la onda P y el comienzo del complejo QRS es el intervalo que hay entre el inicio de la excitación eléctrica de las aurículas y el inicio de la excitación de los ventrículos. Este período se denomina intervalo P-Q. El intervalo P-Q normal es de aproximadamente 0,16 s. (Con frecuencia este intervalo se denomina intervalo P-R porque es probable que no haya onda Q.) Intervalo Q-T La contracción del ventrículo dura casi desde el comienzo de la onda Q (onda R si no hay onda Q) hasta el final de la onda T. Este intervalo se denomina intervalo Q-T y habitualmente es de aproximadamente 0,35 s.
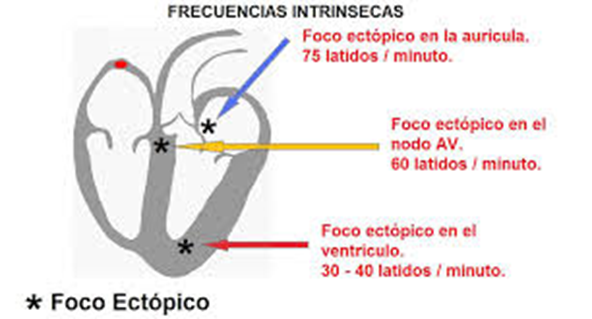
Determinación de la frecuencia del latido cardíaco a partir del electrocardiograma.
La frecuencia del latido cardíaco se puede determinar fácilmente a partir del ECG porque la frecuencia cardíaca es el recíproco del intervalo de tiempo entre dos latidos cardíacos sucesivos. Si el intervalo entre dos latidos, que se determina a partir de las líneas de calibración del tiempo, es de 1 s, la frecuencia cardíaca es de 60 latidos/min. El intervalo normal entre dos complejos QRS sucesivos en una persona adulta es de aproximadamente 0,83 s, lo que corresponde a una frecuencia cardíaca de 60/0,83 veces por minuto, o 72 latidos/mi.
Flujo de corriente alrededor del corazón durante el ciclo cardíaco Registro de potenciales eléctricos a partir de una masa parcialmente despolarizada de músculo cardíaco sincitial
Flujo de corrientes eléctricas en el tórax alrededor del corazón La imagen inferior muestra el músculo ventricular situado en el interior del tórax. Incluso los pulmones, aunque están llenos de aire en su mayor parte, conducen la electricidad en una magnitud sorprendente, y los líquidos de los demás tejidos que rodean el corazón conducen la electricidad incluso con más facilidad. Por tanto, el corazón realmente está suspendido en un medio conductor. Cuando una porción de los ventrículos se despolariza y, por tanto, se hace electronegativa en relación con el resto, la corriente eléctrica fluye desde la zona despolarizada hacia la zona polarizada en rutas sinuosas largas
la primera zona de los ventrículos a la que llega el impulso cardíaco es el tabique, y poco después se propaga a la superficie interna del resto de la masa de los ventrículos, como se muestra por las zonas rojas y los signos negativos.
Este proceso hace que las zonas internas de los ventrículos sean electronegativas y que las paredes externas de los ventrículos sean electropositivas, de modo que la corriente eléctrica fluye a través de los líquidos que rodean los ventrículos en trayectos elípticos, como señalan las flechas curvas de la figura. Si se realiza el promedio algebraico de todas las líneas de flujo de corriente (las líneas elípticas) se encuentra que el flujo medio de corriente tiene negatividad hacia la base del corazón y positividad hacia la punta. Durante la mayor parte del resto del proceso de despolarización la corriente también sigue fluyendo en esta misma dirección, mientras que la despolarización se propaga desde la superficie endocárdica hacia el exterior a través de la masa del músculo ventricular. Después, inmediatamente antes de que la despolarización haya completado su trayecto a través de los ventrículos, la dirección media del flujo de corriente se invierte durante aproximadamente 0,01 s, fluyendo desde la punta ventricular hacia la base, porque la última parte del corazón que se despolariza son las paredes externas de los ventrículos cerca de la base del corazón. Así, en los ventrículos del corazón normal la corriente fluye desde las zonas negativas a las positivas principalmente en una dirección que va desde la base del corazón hacia la punta durante casi todo el ciclo de despolarización, excepto al final.
Derivaciones electrocardiográficas
Tres derivaciones bipolares de las extremidades
las conexiones eléctricas entre las extremidades del paciente y el electrocardiógrafo para registrar ECG de las denominadas derivaciones bipolares estándar de las extremidades. El término «bipolar» significa que el electrocardiograma se registra a partir de dos electrodos que están localizados en lados diferentes del corazón, en este caso en las extremidades. Así, una «derivación» no es un único cable que procede del cuerpo, sino una combinación de dos cables y sus electrodos para formar un circuito completo entre el cuerpo y el electrocardiógrafo. En cada uno de los casos el electrocardiógrafo se representa en el diagrama mediante un medidor eléctrico, aunque el electrocardiógrafo real es un sistema informático de alta velocidad con una pantalla electrónica.
Derivación I Cuando se registra la derivación I, el terminal negativo del electrocardiógrafo está conectado al brazo derecho y el terminal positivo al brazo izquierdo. Por tanto, cuando el punto en el que el brazo derecho se conecta con el tórax es electronegativo respecto al punto en el que se conecta el brazo izquierdo el electrocardiógrafo registra una señal positiva, es decir, por encima de la línea de voltaje cero del ECG. Cuando ocurre lo contrario el electrocardiógrafo registra una señal por debajo de la línea.
Derivación II Para registrar la derivación II de las extremidades, el terminal negativo del electrocardiógrafo se conecta al brazo derecho y el terminal positivo a la pierna izquierda. Por tanto, cuando el brazo derecho es negativo respecto a la pierna izquierda, el electrocardiógrafo registra una señal positiva.
Derivación III Para registrar la derivación III de las extremidades, el terminal negativo del electrocardiógrafo se conecta al brazo izquierdo y el terminal positivo a la pierna izquierda. Esta configuración significa que el electrocardiógrafo registra una señal positiva cuando el brazo izquierdo es negativo respecto a la pierna izquierda.
Triángulo de Einthoven Se dibuja un triángulo, denominado triángulo de Einthoven, alrededor de la zona del corazón. Este diagrama ilustra que los dos brazos y la pierna izquierda forman vértices de un triángulo que rodea el corazón. Los dos vértices de la parte superior del triángulo representan los puntos en los que los dos brazos se conectan eléctricamente a los líquidos que rodean el corazón y el vértice izquierdo es el punto en el que la pierna izquierda se conecta a los líquidos.
Ley de Einthoven La ley de Einthoven afirma que si los ECG se registran simultáneamente en las tres derivaciones de las extremidades, la suma de los potenciales registrados en las derivaciones I y III debe ser igual al potencial en la derivación II. En otras palabras, si en cualquier momento dado se conocen los potenciales eléctricos de dos cualesquiera de las tres derivaciones electrocardiográficas bipolares de las extremidades, se puede determinar la tercera simplemente sumando las dos primeras. Ha de tenerse en cuenta, sin embargo, que se deben observar los signos positivos y negativos de las diferentes derivaciones cuando se haga esta suma.
Derivaciones del tórax (derivaciones precordiales) Con frecuencia se registran ECG con un electrodo situado en la superficie anterior del tórax directamente sobre el corazón Este electrodo se conecta al terminal positivo del electrocardiógrafo, y el electrodo negativo, denominado electrodo indiferente, se conecta a través de resistencias eléctricas iguales al brazo derecho, al brazo izquierdo y a la pierna izquierda al mismo tiempo, como también se muestra en la figura. Habitualmente se registran seis derivaciones estándar del tórax, una cada vez, desde la pared torácica anterior, de modo que el electrodo del tórax se coloca secuencialmente en los seis puntos que se muestran en el diagrama. Los diferentes registros se conocen como derivaciones V1, V2, V3, V4, V5 y V6.
Derivaciones unipolares ampliadas de las extremidades Otro sistema de derivaciones que se utiliza mucho es la derivación unipolar ampliada de las extremidades. En este tipo de registro, dos de las extremidades se conectan mediante resistencias eléctricas al terminal negativo del electrocardiógrafo, y la tercera extremidad se conecta al terminal positivo. Cuando el terminal positivo está en el brazo derecho la derivación se conoce como derivación aVR, cuando está en el brazo izquierdo es la derivación aVL y cuando está en la pierna izquierda es la derivación aVF.
El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto porque la duración de la contracción es mucho mayor.
el músculo cardíaco tiene las miofibrillas típicas que contienen filamentos de actina y de miosina casi idénticos a los que se encuentran en el músculo esquelético; estos filamentos están unos al lado de otros y se deslizan durante la contracción de la misma manera que ocurre en el músculo esquelético.
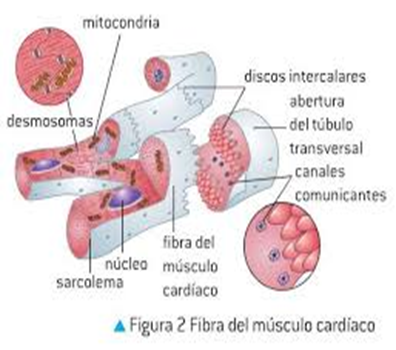
músculo cardíaco es un sincitio:
Las zonas oscuras que atraviesan las fibras musculares cardíacas se le denominan discos intercalados, las fibras musculares cardíacas están formadas por muchas células individuales conectadas entre sí en serie y en paralelo.
En cada uno de los discos intercalados las membranas celulares se fusionan entre sí para formar uniones «comunicantes» (en hendidura) permeables que permiten una rápida difusión. desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular a lo largo del eje longitudinal de las fibras musculares cardíacas, de modo que los potenciales de acción viajan fácilmente desde una célula muscular cardíaca a la siguiente, a través de los discos intercalados. Por tanto, el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una célula se excita el potencial de acción se propaga rápidamente a todas.
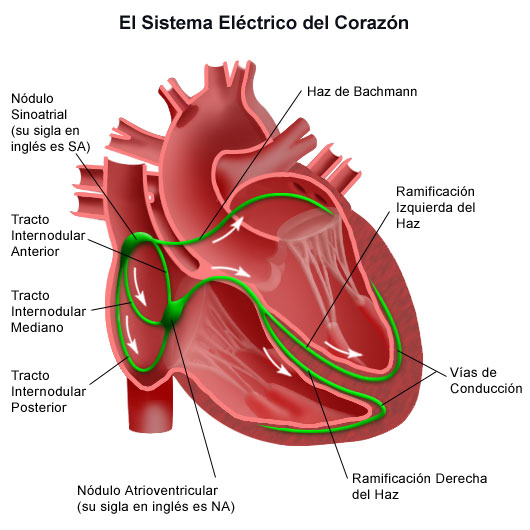
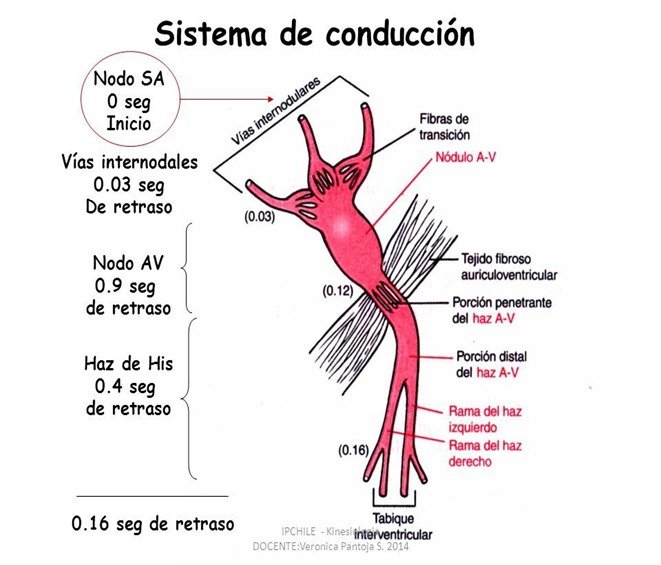
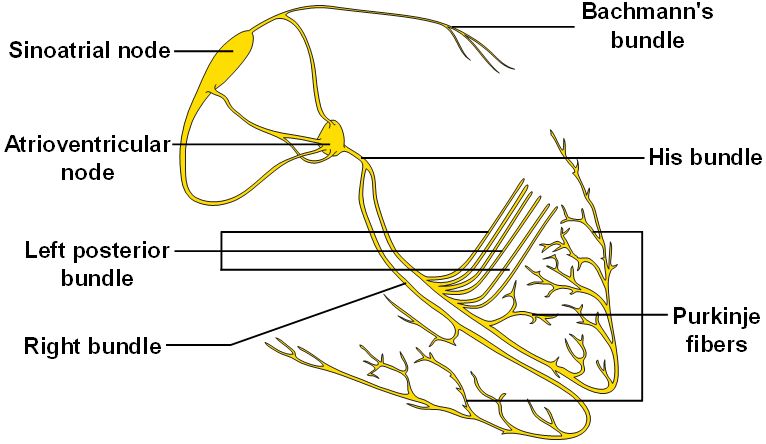
El corazón realmente está formado por dos sincitios: el sincitio auricular, que forma las paredes de las dos aurículas, y el sincitio ventricular, que forma las paredes de los dos ventrículos. Las aurículas están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares (AV) entre las aurículas y los ventrículos. Normalmente los potenciales no se conducen desde el sincitio auricular hacia el sincitio ventricular directamente a través de este tejido fibroso. Por el contrario, solo son conducidos por medio de un sistema de conducción especializado denominado haz AV, que es un fascículo de fibras de conducción de varios milímetros de diámetro.
Potenciales de acción en el músculo cardíaco:
El potencial de acción que se registra en una fibra muscular ventricular, es en promedio de aproximadamente 105 mV, lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproximadamente –85 mV, entre los latidos hasta un valor ligeramente positivo, de aproximadamente +20 mV, durante cada latido. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta, seguida al final de la meseta de una repolarización súbita. La presencia de esta meseta del potencial de acción ha- ce que la contracción ventricular dure hasta 15 veces más en el músculo cardíaco que en el músculo esquelético.
el potencial de acción del músculo esquelético está producido casi por completo por la apertura súbita de grandes números de canales rápidos de sodio que permiten que grandes cantidades de iones sodio entren en la fibra muscular esquelética desde el líquido extracelular. A estos canales se les denomina canales «rápidos» porque permanecen abiertos solo algunas milésimas de segundo y después se cierran súbitamente. Al final de este cierre se produce la repolarización y el potencial de acción ha terminado en otra milésima de segundo aproximadamente
el potencial de acción está producido por la apertura de dos tipos de canales: 1) los mismos canales rápidos de sodio activados por el voltaje que en el músculo. esquelético y 2) otra población totalmente distinta de canales de calcio de tipo L (canales lentos de calcio), que también se denominan canales de calcio-sodio.
La segunda diferencia funcional importante entre el músculo cardíaco y el músculo esquelético que ayuda a explicar tanto el potencial de acción prolongado como su meseta es la siguiente: inmediatamente después del inicio del potencial de acción, la permeabilidad de la membrana del músculo cardíaco a los iones potasio disminuye aproximadamente cinco veces, un efecto que no aparece en el músculo esquelético.
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de unos 0,3 a 0,5 m/s, o aproximadamente 1/250 de la velocidad en las fibras nerviosas grandes y en torno a 1/10 de la velocidad en las fibras musculares esqueléticas. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje, es de hasta 4 m/s en la mayoría de las partes del sistema, lo que permite una conducción razonablemente rápida de la señal excitadora hacia las diferentes partes del corazón.
Duración de la contracción El músculo cardíaco comienza a contraerse algunos milisegundos después de la llegada del potencial de acción y sigue contrayéndose hasta algunos milisegundos después de que finalice. la duración de la contracción del músculo cardíaco depende principalmente de la duración del potencial de acción, incluyendo la meseta, aproximadamente 0,2 s en el músculo auricular y 0,3 s en el músculo ventricular.
Diástole y sístole El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole. La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca. Por ejemplo, si la frecuencia cardíaca es de 72 latidos/min, la duración del ciclo cardíaco es de 1/72 min/latido, aproximadamente 0,0139 min por latido, o 0,833 s por latido. los diferentes acontecimientos que se producen durante el ciclo cardíaco para el lado izquierdo del corazón. Las tres curvas superiores muestran los cambios de presión en la aorta, en el ventrículo izquierdo y en la aurícula izquierda, respectivamente. La cuarta curva representa los cambios del volumen ventricular izquierdo, la quinta el electrocardiograma y la sexta un fonocardiograma, que es un registro de los ruidos que produce el corazón (principalmente las válvulas cardíacas) durante su función de bombeo.
La sangre normalmente fluye de forma continua desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas. Después, la contracción auricular habitualmente produce un llenado de un 20% adicional de los ventrículos. Por tanto, las aurículas actúan como bombas de cebado que aumentan la eficacia del bombeo ventricular hasta un 20%. el corazón puede seguir funcionando en la mayor parte de las condiciones incluso sin esta eficacia de un 20% adicional porque normalmente tiene la capacidad de bombear entre el 300 y el 400% más de sangre de la que necesita el cuerpo en reposo. Por tanto, cuando las aurículas dejan de funcionar es poco probable que se observe esta diferencia salvo que la persona haga un esfuerzo; en este caso de manera ocasional aparecen síntomas agudos de insuficiencia cardíaca, especialmente disnea.
Los ventrículos se llenan de sangre durante la diástole:
Durante la sístole ventricular se acumulan grandes cantidades de sangre en las aurículas derecha e izquierda porque las válvulas AV están cerradas. Por tanto, tan pronto como ha finalizado la sístole y las presiones ventriculares disminuyen de nuevo a sus valores diastólicos bajos, el aumento moderado de presión que se ha generado en las aurículas durante la sístole ventricular inmediatamente abre las válvulas AV y permite que la sangre fluya rápidamente hacia los ventrículos, como se muestra en la elevación de la curva de volumen ventricular izquierdo. Es el denominado período de llenado rápido de los ventrículos. El período de llenado rápido dura aproximadamente el primer tercio de la diástole. Durante el tercio medio de la diástole normalmente solo fluye una pequeña cantidad de sangre hacia los ventrículos; esta es la sangre que continúa drenando hacia las aurículas desde las venas y que pasa a través de las aurículas directamente hacia los ventrículos. Durante el último tercio de la diástole las aurículas se contraen y aportan un impulso adicional al flujo de entrada de sangre hacia los ventrículos.
Válvulas auriculoventriculares
Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmona hacia los ventrículos durante la diástole. para el ventrículo izquierdo, se cierran y abren pasivamente. Es decir, se cierran cuando un gradiente de presión retrógrada empuja la sangre hacia atrás, y se abren cuando un gradiente de presión anterógrada fuerza la sangre en dirección anterógrada. Por motivos anatómicos, las válvulas AV, que están formadas por una película delgada, casi no precisan ningún flujo retrógrado para cerrarse, mientras que las válvulas semilunares, que son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
Los músculos papilares
se contraen cuando se contraen las paredes ventriculares, pero, al contrario de lo que se podría esperar, no contribuyen al cierre de las válvulas. Por el contrario, tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular. Si se produce la rotura de una cuerda tendinosa o si se produce parálisis de uno de los músculos papilares, la válvula protruye mucho hacia las aurículas durante la contracción ventricular, a veces tanto que se produce una fuga grave y da lugar a una insuficiencia cardíaca grave o incluso mortal.
Válvulas aórtica y de la arteria pulmonar Las válvulas semilunares aórtica y pulmonar funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente, a diferencia del cierre mucho más suave de las válvulas AV. Segundo, debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores. Además, debido al cierre rápido y a la eyección rápida, los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV. Las válvulas AV tienen el soporte de las cuerdas tendinosas, lo que no ocurre en el caso de las válvulas semilunares. A partir de la anatomía de las válvulas aórtica y pulmonar (que se muestra para la válvula aórtica en la parte inferior de la figura 9-8) es evidente que deben estar situadas sobre una base de un tejido fibroso especialmente fuerte, pero muy flexible para soportar las tensiones físicas adicionales.
Válvulas aórtica y de la arteria pulmonar
Las válvulas semilunares aórtica y pulmonar
funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente, a diferencia del cierre mucho más suave de las válvulas AV Segundo, debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores. Además, debido al cierre rápido y a la eyección rápida, los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV. Las válvulas AV tienen el soporte de las cuerdas tendinosas, lo que no ocurre en el caso de las válvulas semilunares. A partir de la anatomía de las válvulas aórtica y pulmonar (que se muestra para la válvula aórtica en la parte inferior, es evidente que deben estar situadas sobre una base de un tejido fibroso especialmente fuerte, pero muy flexible para soportar las tensiones físicas adicionales.
Regulación del bombeo cardíaco Cuando una persona está en reposo el corazón solo bombea de 4 a 6 l de sangre cada minuto. Durante el ejercicio intenso puede ser necesario que el corazón bombee de cuatro a siete veces esta cantidad. Los mecanismos básicos mediante los que se regula el volumen que bombea el corazón son:
1)regulación cardíaca intrínseca del bombeo en respuesta a los cambios del volumen de la sangre que fluye hacia el corazón
2) control de la frecuencia cardíaca y del bombeo cardíaco por el sistema nervioso autónomo.
Mecanismo de Frank-Starling:
La capacidad intrínseca del corazón de adaptarse a volúmenes crecientes de flujo sanguíneo de entrada se denomina mecanismo de Frank-Starling del corazón en honor de Otto Frank y Ernest Starling, dos grandes fisiólogos de hace un siglo. Básicamente, el mecanismo de Frank-Starling significa que cuanto más se distiende el músculo cardíaco durante el llenado, mayor es la fuerza de contracción y mayor es la cantidad de sangre que bombea hacia la aorta. O, enunciado de otra manera, dentro de límites fisiológicos el corazón bombea toda la sangre que le llega procedente de las venas.
Cuando una cantidad adicional de sangre fluye hacia los ventrículos, el propio músculo cardíaco es distendido hasta una mayor longitud. Esta distensión, a su vez, hace que el músculo se contraiga con más fuerza porque los filamentos de actina y de miosina son desplazados hacia un grado más óptimo de superposición para la generación de fuerza. Por tanto, el ventrículo, debido al aumento de la función de bomba, bombea automáticamente la sangre adicional hacia las arterias. Esta capacidad del músculo distendido, hasta una longitud óptima, de contraerse con un aumento del trabajo cardíaco, es característica de todo el músculo estriado. no es simplemente una característica del músculo cardíaco. Es importante efecto del aumento de longitud del músculo cardíaco, hay otro factor que aumenta la función de bomba del corazón cuando aumenta su volumen. La distensión de la pared de la aurícula derecha aumenta directamente la frecuencia cardíaca en un 10-20%, lo que también contribuye a aumentar la cantidad de sangre que se bombea cada minuto, aunque su contribución es mucho menor que la del mecanismo de Frank-Starling.
Control del corazón por los nervios simpáticos y parasimpáticos
La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos y parasimpáticos (vagos), que inervan de forma abundante el corazón.
Para niveles dados de presión auricular de entrada, la cantidad de sangre que se bombea cada minuto (gasto cardíaco) con frecuencia se puede aumentar más de un 100% por la estimulación simpática. Por el contrario, el gasto se puede disminuir hasta un valor tan bajo como cero o casi cero por la estimulación vagal (parasimpática).
La estimulación simpática intensa puede aumentar la frecuencia cardíaca en seres humanos adultos jóvenes desde la frecuencia normal de 70 latidos/min hasta 180 a 200 y, raras veces, incluso 250 latidos/min. Además, la estimulación simpática aumenta la fuerza de la contracción cardíaca hasta el doble de lo normal, aumentando de esta manera el volumen de sangre que se bombea y aumentando la presión de eyección. Así, con frecuencia la estimulación simpática puede aumentar el gasto cardíaco máximo hasta dos o tres veces, además del aumento del gasto que produce el mecanismo de Frank-Starling que ya se ha comentado. Por el contrario, la inhibición de los nervios simpáticos del corazón puede disminuir la función de bomba del corazón en un grado moderado: en condiciones normales, las fibras nerviosas simpáticas que llegan al corazón descargan continuamente a una frecuencia baja que mantiene el bombeo aproximadamente un 30% por encima del que habría sin estimulación simpática. Por tanto, cuando la actividad del sistema nervioso simpático disminuye por debajo de lo normal, tanto de la frecuencia cardíaca como la fuerza de la contracción del músculo ventricular se reducen, con lo que disminuye el nivel de bombeo cardíaco hasta un 30% por debajo de lo normal.
La estimulación parasimpática (vagal) reduce la frecuencia cardíaca y la fuerza de la contracción
La estimulación intensa de las fibras nerviosas parasimpáticas de los nervios vagos que llegan al corazón puede interrumpir el latido cardíaco durante algunos segundos, pero después el corazón habitualmente «escapa» y late a una frecuencia de 20 a 40 latidos/min mientras continúe la estimulación parasimpática. Además, la estimulación vagal intensa puede reducir la fuerza de la contracción del músculo cardíaco en un 20-30%. Las fibras vagales se distribuyen principalmente por las aurículas y no mucho en los ventrículos, en los que se produce la contracción de potencia del corazón. Esta distribución explica por qué el efecto de la estimulación vagal tiene lugar principalmente sobre la reducción de la frecuencia cardíaca, en lugar de reducir mucho la fuerza de la contracción del corazón. Sin embargo, la gran disminución de la frecuencia cardíaca, combinada con una ligera reducción de la fuerza de la contracción cardíaca, puede reducir el bombeo ventricular en un 50% o más.
Efecto de los iones potasio
El exceso de potasio hace que el corazón esté dilatado y flácido, y también reduce la frecuencia cardíaca. Grandes cantidades de potasio también pueden bloquear la conducción del impulso cardíaco desde las aurículas hacia los ventrículos a través del haz AV. La elevación de la concentración de potasio hasta solo 8 a 12 mEq/l (dos a tres veces el valor normal) puede producir una profunda debilidad del corazón, una alteración del ritmo e incluso la muerte. Estos efectos se deben parcialmente al hecho de que una concentración elevada de potasio en los líquidos extracelulares reduce el potencial de membrana en reposo de las fibras del músculo cardíaco, como se explica en el capítulo 5. Es decir, la alta concentración de potasio en el líquido extracelular despolariza parcialmente la membrana celular, lo que provoca que el potencial de membrana sea menos negativo. Cuando disminuye el potencial de membrana también lo hace la intensidad del potencial de acción, lo que hace que la contracción del corazón sea progresivamente más débil.
Efecto de los iones calcio
Un exceso de iones calcio produce efectos casi exactamente contrarios a los de los iones potasio, haciendo que el corazón progrese hacia una contracción espástica. Este efecto está producido por el efecto directo de los iones calcio en el inicio del proceso contráctil cardíaco, como se explicó antes en este mismo capítulo. Por el contrario, el déficit de iones calcio produce debilidad cardíaca, similar al efecto de la elevación de la concentración de potasio. Afortunadamente las concentraciones de iones calcio en la sangre normalmente están reguladas en un intervalo muy estrecho. Por tanto, los efectos cardíacos de las concentraciones anormales de calcio raras veces tienen significado clínico.
Efecto de la temperatura sobre la función cardíaca
El aumento de la temperatura corporal, como ocurre durante la fiebre, produce un gran aumento de la frecuencia cardíaca, a veces hasta del doble del valor normal. El descenso de la temperatura produce una gran disminución de la frecuencia cardíaca, que puede disminuir hasta solo algunos latidos por minuto cuando una persona está cerca de la muerte por hipotermia en el intervalo de temperatura corporal de 16 °C a 21 °C. Estos efectos probablemente se deben al hecho de que el calor aumenta la permeabilidad de la membrana del músculo cardíaco a los iones que controlan la frecuencia cardíaca, acelerando el proceso de autoexcitación. La fuerza contráctil del corazón con frecuencia se incrementa transitoriamente cuando hay un aumento moderado de la temperatura, como ocurre durante el ejercicio corporal, aunque una elevación prolongada de la temperatura agota los sistemas metabólicos del corazón y finalmente produce debilidad. Por tanto, la función óptima del corazón depende mucho del control adecuado de la temperatura corporal mediante los mecanismos
El corazón humano tiene un sistema especial para la autoexcitación rítmica y la contracción repetitiva aproximadamente 100.000 veces al día.
Sistema de excitación especializado y de conducción del corazón
el nódulo sinusal (también denominado nódulo sinoauricular o SA), en el que se genera el impulso rítmico normal; las vías internodulares que conducen impulsos desde el nódulo sinusal hasta el nódulo auriculoventricular (AV); el nódulo AV, en el cual los impulsos originados en las aurículas se retrasan antes de penetrar en los ventrículos; el haz AV, que conduce impulsos desde las aurículas hacia los ventrículos, y las ramas izquierda y derecha del haz de fibras de Purkinje, que conducen los impulsos cardíacos por todo el tejido de los ventrículos.
Nódulo sinusal (sinoauricular) El nódulo sinusal (también denominado nódulo sinoauricular) es una banda elipsoide, aplanada y pequeña de músculo cardíaco especializado de aproximadamente 3 mm de anchura, 15 mm de longitud y 1 mm de grosor. Está localizado en la pared posterolateral superior de la aurícula derecha.
Las fibras de este nódulo casi no tienen filamentos musculares contráctiles y cada una de ellas tiene solo de 3 a 5 μm de diámetro, sus fibras se conectan directamente con las fibras musculares auriculares, de modo que todos los potenciales de acción que comienzan en el nódulo sinusal se propagan inmediatamente hacia la pared del músculo auricular.
Ritmicidad eléctrica automática de las fibras sinusales
La autoexcitación es un proceso que puede producir descargas y contracciones rítmicas automáticas. Esta capacidad es cierta en el caso de las fibras del sistema especializado de conducción del corazón.
Mecanismo de la ritmicidad del nódulo sinusal
El «potencial de membrana en reposo» de la fibra del nódulo sinusal tiene una negatividad de aproximadamente –55 a –60 mV, en comparación con –85 a –90 mV para la fibra muscular ventricular. La causa de esta menor negatividad es que las membranas celulares de las fibras sinusales son permeables naturalmente a los iones sodio y calcio, y las cargas positivas de los iones sodio y calcio que entran neutralizan parte de la negatividad intracelular.
El músculo cardíaco tiene tres tipos de canales iónicos de membrana que tienen funciones importantes en la generación de los cambios de voltaje en el potencial de acción. Los tipos son: 1) los canales rápidos de sodio; 2) los canales de calcio de tipo L (canales lentos de sodio-calcio), y 3) los canales de potasio. La apertura de los canales rápidos de sodio durante algunas diezmilésimas de segundo es responsable de la rápida espiga ascendente del potencial de acción que se observa en el músculo ventricular, debido a la entrada rápida de iones sodio positivos hacia el interior de la fibra. Después, la «meseta» del potencial de acción ventricular está producida principalmente por la apertura más lenta de los canales lentos de sodio-calcio, que dura aproximadamente 0,3 s. Finalmente, la apertura de los canales de potasio permite la difusión de grandes cantidades de iones potasio positivos hacia el exterior a través de la membrana de la fibra y devuelve el potencial de membrana a su nivel de reposo.
Autoexcitación de las fibras del nódulo sinusal
Los iones sodio positivos del exterior de las fibras normalmente tienden a desplazarse hacia el interior. Entre los latidos cardíacos, la entrada de iones sodio de carga positiva produce una elevación lenta del potencial de membrana en reposo en dirección positiva.
El potencial «en reposo» aumenta gradualmente y se hace menos negativo entre cada dos latidos sucesivos. Cuando el potencial alcanza un voltaje umbral de aproximadamente –40 mV, los canales de calcio de tipo L se «activan», produciendo de esta manera el potencial de acción.
Durante el transcurso del potencial de acción se producen dos fenómenos que impiden dicho estado de despolarización constante. Primero, los canales de calcio de tipo L se inactivan en un plazo de aproximadamente 100 a 150 ms después de su apertura, y segundo, aproximadamente al mismo tiempo se abren números muy elevados de canales de potasio. Por tanto, se interrumpe el flujo de entrada de iones positivos calcio y sodio a través de los canales de calcio de tipo L, mientras que al mismo tiempo grandes cantidades de iones positivos de potasio difunden hacia el exterior de la fibra. ponen fin al potencial de acción. Además, los canales de potasio permanecen abiertos durante algunas décimas de segundo más, manteniendo transitoriamente el movimiento de cargas positivas hacia el exterior de la célula, con el consiguiente exceso de negatividad en el interior de la fibra; este proceso se denomina hiperpolarización.
El estado de hiperpolarización inicialmente desplaza el potencial de membrana «en reposo» hacia abajo hasta aproximadamente –55 o –60 mV al final del potencial de acción.
Las vías internodulares e interauriculares transmiten impulsos cardíacos a través de las aurículas.
Los extremos de las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares circundantes. Los potenciales de acción que se originan en el nódulo sinusal viajan hacia estas fibras musculares auriculares. De esta manera, el potencial de acción se propaga por toda la masa muscular auricular y, finalmente, llega hasta el nódulo AV. La velocidad de conducción en la mayor parte del músculo auricular es de aproximadamente 0,3 m/s.
La banda interauricular anterior, atraviesa las paredes anteriores de las aurículas para dirigirse hacia la aurícula izquierda. Además, otras tres bandas pequeñas se incurvan a través de las paredes auriculares anterior, lateral y posterior, y terminan en el nódulo AV
El nódulo auriculoventricular retrasa la conducción del impulso desde las aurículas a los ventrículos
Existe un retraso de tiempo para que las aurículas vacíen su sangre hacia los ventrículos antes de que comience la contracción ventricular.
El retraso de la transmisión hacia los ventrículos se produce principalmente en el nódulo AV y en sus fibras de conducción adyacentes.
Causa de la conducción lenta La conducción lenta en las fibras transicionales, nodulares y penetrantes del haz AV está producida principalmente por la disminución del número de uniones en hendidura entre células sucesivas de las vías de conducción, de modo que hay una gran resistencia a la conducción de los iones excitadores desde una fibra de conducción hasta la siguiente.
Transmisión rápida en el sistema de Purkinje ventricular Las fibras de Purkinje especiales se dirigen desde el nódulo AV a través del haz AV hacia los ventrículos. Excepto en la porción inicial de estas fibras, donde penetran en la barrera fibrosa AV, tienen características funcionales bastante distintas a las de las fibras del nódulo AV. Son fibras muy grandes, incluso mayores que las fibras musculares ventriculares normales, y transmiten potenciales de acción a una velocidad de 1,5 a 4 m/s, una velocidad aproximadamente seis veces mayor que la del músculo ventricular normal y 150 veces mayor que la de algunas de las fibras del nódulo AV. Esta velocidad permite una transmisión casi instantánea del impulso cardíaco por todo el resto del músculo ventricular. Las fibras de Purkinje también tienen muy pocas miofibrillas, lo que significa que se contraen poco o nada durante la transmisión de los impulsos.
Conducción unidireccional a través del haz AV
Una característica especial del haz AV es la imposibilidad, excepto en estados anormales, de que los potenciales de acción viajen retrógradamente desde los ventrículos hacia las aurículas. Esta característica impide la reentrada de los impulsos cardíacos por esta ruta desde los ventrículos hacia las aurículas, permitiendo solo la contracción anterógrada desde las aurículas hacia los ventrículos.
El haz AV el músculo auricular está separado del músculo ventricular por una barrera fibrosa. Esta barrera normalmente actúa como aislante para impedir el paso de los impulsos cardíacos entre el músculo auricular y ventricular a través de cualquier ruta distinta a la conducción anterógrada a través del propio haz AV.
Distribución de las fibras de Purkinje en los ventrículos: las ramas izquierda y derecha del haz
Después de penetrar en el tejido fibroso que está entre el músculo auricular y ventricular, la porción distal del haz AV se dirigía hacia abajo en el interior del tabique interventricular a lo largo de 5 a 15 mm hacia la punta del corazón. Después el haz se divide en las ramas izquierda y derecha del haz, que están debajo del endocardio en los dos lados respectivos del tabique interventricular.
Cada una de las ramas se dirige hacia abajo, hacia la punta del ventrículo, dividiéndose progresivamente en ramas más pequeñas. Estas ramas, a su vez, siguen un trayecto en dirección lateral alrededor de cada una de las cavidades ventriculares y hacia atrás, hacia la base del corazón. Las fibras de Purkinje penetran en aproximadamente un tercio del grosor de la masa muscular y finalmente se continúan con las fibras musculares cardíacas. Desde el momento en el que el impulso cardíaco entre las ramas del haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de Purkinje el tiempo total transcurrido es en promedio de sólo 0,03 s.
Transmisión del impulso cardíaco en el músculo ventricular
Una vez que el impulso llega a los extremos de las fibras de Purkinje se transmite a través de la masa del músculo ventricular por las propias fibras musculares ventriculares. La velocidad de transmisión es ahora solo de 0,3 a 0,5 m/s, una sexta parte de la velocidad de las fibras de Purkinje.
Así, el tiempo total para la transmisión del impulso cardíaco desde las ramas iniciales del haz hasta las últimas fibras del músculo ventricular en el corazón normal es de aproximadamente 0,06 s.
Control de la excitación y la conducción en el corazón
El nódulo sinusal es el marcapasos normal del corazón.
Las fibras del nódulo AV, cuando no son estimuladas por alguna fuente externa, descargan a una frecuencia rítmica intrínseca de 40 a 60 veces por minuto, y las fibras de Purkinje lo hacen a una frecuencia de entre 15 y 40 veces por minuto. Estas frecuencias son distintas a la frecuencia normal del nódulo sinusal, de 70 a 80 veces por minuto.
Cada vez que se produce una descarga en el nódulo sinusal su impulso se conduce hacia el nódulo AV y hacia las fibras de Purkinje, produciendo también la descarga de sus membranas. El impulso procedente del nódulo sinusal descarga tanto las fibras del nódulo AV como las fibras de Purkinje antes de que se pueda producir autoexcitación en cualquiera de esas estructuras.
Marcapasos anormales: marcapasos «ectópico»
Un marcapasos que está situado en una localización distinta al nódulo sinusal se denomina marcapasos «ectópico». Un marcapasos ectópico da lugar a una secuencia anormal de contracción de las diferentes partes del corazón y puede producir una debilidad significativa del bombeo cardíaco. El nuevo marcapasos se produce en este caso con más frecuencia en el nódulo AV o en la porción penetrante del haz AV en su trayecto hacia los ventrículos.
cuando el impulso cardíaco no puede pasar desde las aurículas hacia los ventrículos a través del sistema del nódulo AV y del haz, las aurículas siguen latiendo a la frecuencia normal del ritmo del nódulo sinusal, mientras que habitualmente aparece un nuevo marcapasos en el sistema de Purkinje de los ventrículos que activa el músculo ventricular a una frecuencia de entre 15 y 40 latidos/min.
Importancia del sistema de Purkinje en la generación de una contracción sincrónica del músculo ventricular
La rápida conducción del sistema de Purkinje permite normalmente que el impulso cardíaco llegue a casi todas las porciones de los ventrículos en un breve intervalo de tiempo, excitando la primera fibra muscular ventricular solo 0,03 a 0,06 s antes de la excitación de la última. Esta sincronización hace que todas las porciones del músculo de los dos ventrículos comienzan a contraerse casi al mismo tiempo y que después sigan contrayéndose durante aproximadamente otros 0,3 s.
Los nervios simpáticos y parasimpáticos controlan el ritmo cardíaco y la conducción de impulsos por los nervios cardíacos
Los nervios parasimpáticos (vagos) se distribuyen principalmente a los nódulos SA y AV, en mucho menor grado al músculo de las dos aurículas y apenas directamente al músculo ventricular. Por el contrario, los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular, así como en todas las demás zonas.
La estimulación parasimpática (vagal) ralentiza el ritmo y la conducción cardíacos.
La estimulación de los nervios parasimpáticos que llegan al corazón (los vagos) hace que se libere la hormona acetilcolina en las terminaciones nerviosas. Esta hormona tiene dos efectos principales sobre el corazón. Primero, reduce la frecuencia del ritmo del nódulo sinusal, y segundo, reduce la excitabilidad de las fibras de la unión AV entre la musculatura auricular y el nódulo AV, retrasando de esta manera la transmisión del impulso cardíaco hacia los ventrículos. Una estimulación vagal débil a moderada reduce la frecuencia del bombeo del corazón, con frecuencia hasta un valor tan bajo como la mitad de lo normal.
La estimulación intensa de los nervios vagos puede interrumpir completamente la excitación rítmica del nódulo sinusal o puede bloquear completamente la transmisión del impulso cardíaco desde las aurículas hacia los ventrículos a través del nódulo AV. En cualquiera de los casos, las señales excitadores rítmicas ya no se transmiten hacia los ventrículos. Los ventrículos pueden dejar de latir durante 5 a 20 s, pero después alguna área pequeña de las fibras de Purkinje, habitualmente en la porción del tabique interventricular del haz AV, presenta un ritmo propio y genera la contracción ventricular a una frecuencia de 15 a 40 latidos/min. Este fenómeno se denomina escape ventricular.
Mecanismo de los efectos vagales
La acetilcolina que se libera en las terminaciones nerviosas vagales aumenta mucho la permeabilidad de las membranas de las fibras a los iones potasio, lo que permite la salida rápida de potasio desde las fibras del sistema de conducción. Este proceso da lugar a un aumento de la negatividad en el interior de las fibras, un efecto que se denomina hiperpolarización. En el nódulo sinusal, el estado de hiperpolarización hace el potencial de membrana «en reposo» de las fibras del nódulo sinusal mucho más negativo de lo habitual, es decir, de –65 a –75 mV en lugar del nivel normal de –55 a –60 mV.
En el nódulo AV, el estado de hiperpolarización producido por la estimulación vagal hace que sea difícil que las pequeñas fibras auriculares que entran en el nódulo generen una corriente de una intensidad suficiente como para excitar las fibras nodulares. Por tanto, el factor de seguridad para la transmisión del impulso cardíaco a través de las fibras de transición hacia las fibras del nódulo AV disminuye. Una reducción moderada simplemente retrasa la conducción del impulso, aunque una disminución grande bloquea totalmente la conducción.
La estimulación simpática aumenta el ritmo y la conducción del corazón La estimulación simpática produce esencialmente los efectos contrarios sobre el corazón a los que produce la estimulación vagal. Segundo, aumenta la velocidad de conducción, así como el nivel de excitabilidad de todas las porciones del corazón. Tercero, aumenta mucho la fuerza de contracción de toda la musculatura cardíaca, tanto auricular como ventricular. La estimulación simpática aumenta la actividad global del corazón. La estimulación máxima casi puede triplicar la frecuencia del latido cardíaco y puede aumentar la fuerza de la contracción del corazón hasta dos veces.
Mecanismo del efecto simpático La estimulación de los nervios simpáticos libera la hormona noradrenalina en las terminaciones nerviosas simpáticas. La noradrenalina estimula, a su vez, los receptores β1-adrenérgicos, que median en los efectos sobre la frecuencia cardíaca. La estimulación β1-adrenérgica actúa sobre las fibras del músculo cardíaco, aunque se piensa que aumenta la permeabilidad de la membrana de las fibras a los iones sodio y calcio. En el nódulo sinusal, un aumento de la permeabilidad a sodio-calcio genera un potencial en reposo más positivo y también produce un aumento de la velocidad del ascenso del potencial de membrana diastólico hacia el nivel liminal para la autoexcitación, acelerando de esta forma la autoexcitación y, por tanto, aumentando la frecuencia cardíaca. En el nódulo AV y en los haces AV, el aumento de la permeabilidad a sodio-calcio hace que sea más fácil que el potencial de acción excite todas las porciones sucesivas de los haces de las fibras de conducción, disminuyendo de esta manera el tiempo de conducción desde las aurículas hasta los ventrículos. El aumento de la permeabilidad a los iones calcio es responsable al menos en parte del aumento de la fuerza contráctil del músculo cardíaco bajo la influencia de la estimulación simpática, porque los iones calcio tienen una función importante en la excitación del proceso contráctil de las miofibrillas.
Después de que los alvéolos se hayan ventilado con aire limpio, la siguiente fase de la respiración es la difusión del oxígeno (O2) desde los alvéolos hacia la sangre pulmonar y la difusión del dióxido de carbono (CO2) en la dirección opuesta, desde la sangre a los alvéolos. El proceso de difusión es simplemente el movimiento aleatorio de moléculas en todas las direcciones a través de la membrana respiratoria y los líquidos adyacentes.
La diferencia de presión provoca difusión de gases a través de líquidos
Del análisis previo es evidente que cuando la presión parcial de un gas es mayor en una zona que en otra zona, habrá una difusión neta desde la zona de presión elevada hacia la zona de presión baja. Las moléculas de la zona de presión elevada, debido a su mayor número, tienen una mayor probabilidad estadística de moverse de manera aleatoria hacia la zona de baja presión que las moléculas que intentan pasar en la otra dirección.
La difusión neta del gas desde la zona de presión elevada hacia la zona de presión baja es igual al número de moléculas que rebotan en esta dirección anterógrada menos el número que rebota en la dirección contraria, que es proporcional a la diferencia de presiones parciales de gas entre las dos zonas, denominada simplemente diferencia de presión para producir la difusión.
Las composiciones del aire alveolar y el aire atmosférico son diferentes Hay varias razones para estas diferencias. Primero, el aire alveolar es sustituido solo de manera parcial por aire atmosférico en cada respiración. Segundo, el O2 se absorbe constantemente hacia la sangre pulmonar desde el aire pulmonar. Tercero, el CO2 está difundiendo constantemente desde la sangre pulmonar hacia los alvéolos. Y cuarto, el aire atmosférico seco que entra en las vías aéreas es humidificado incluso antes de que llegue a los alvéolos.
Humidificación del aire en las vías aéreas El aire atmosférico está compuesto casi totalmente por nitrógeno y oxígeno; normalmente apenas contiene CO2 y poco vapor de agua. Sin embargo, tan pronto como el aire atmosférico entra en las vías aéreas está expuesto a los líquidos que recubren las superficies respiratorias. Incluso antes de que el aire entre en los alvéolos, se humidifica casi totalmente. La presión parcial de vapor de agua a una temperatura corporal normal de 37 °C es de 47 mmHg, que es, por tanto, la presión parcial de vapor de agua del aire alveolar. Como la presión total en los alvéolos no puede aumentar por encima de la presión atmosférica (760 mmHg a nivel del mar), este vapor de agua simplemente diluye todos los demás gases que están en el aire inspirado.
El aire alveolar se renueva lentamente por el aire atmosférico En promedio la capacidad residual funcional de los pulmones (el volumen de aire que queda en los pulmones al final de una espiración normal) en un hombre mide aproximadamente 2.300 ml. Sin embargo, solo 350 ml de aire nuevo entran en los alvéolos en cada inspiración normal y se espira esta misma cantidad de aire alveolar. Por tanto, el volumen de aire alveolar que es sustituido por aire atmosférico nuevo en cada respiración es de solo 1/7 del total, de modo que son necesarias múltiples inspiraciones para intercambiar la mayor parte del aire alveolar.
Importancia de la sustitución lenta del aire alveolar La sustitución lenta del aire alveolar tiene una importancia particular en la prevención de cambios súbitos de las concentraciones de gases en la sangre. Esto hace que el mecanismo de control respiratorio sea mucho más estable de lo que sería de otro modo, y ayuda a prevenir los aumentos y disminuciones excesivos de la oxigenación tisular, de la concentración tisular de CO2 y del pH tisular cuando se produce una interrupción temporal de la respiración.
Concentración y presión parcial de oxígeno en los alvéolos El oxígeno se absorbe continuamente desde los alvéolos hacia la sangre de los pulmones, y continuamente se respira O2 nuevo hacia los alvéolos desde la atmósfera. Cuanto más rápidamente se absorba el O2, menor será su concentración en los alvéolos; por el contrario, cuanto más rápidamente se inhale nuevo O2 hacia los alvéolos desde la atmósfera, mayor será su concentración. Por tanto, la concentración de O2 en los alvéolos, y también su presión parcial, está controlada por: 1) la velocidad de absorción de O2 hacia la sangre
2) la velocidad de entrada de O2 nuevo a los pulmones por el proceso ventilatorio.
Difusión de gases a través de la membrana respiratoria Unidad respiratoria La unidad respiratoria (también denominada «lobulillo respiratorio»), que está formada por un bronquíolo respiratorio, los conductos alveolares, los atrios y los alvéolos.
Hay aproximadamente 300 millones de alvéolos en los dos pulmones, y cada alvéolo tiene un diámetro medio de aproximadamente 0,2 mm. Las paredes alveolares son muy delgadas y entre los alvéolos hay una red casi sólida de capilares interconectados. debido a lo extenso del plexo capilar, se ha descrito que el flujo de sangre en la pared alveolar es una «lámina» de sangre que fluye. Así, es evidente que los gases alveolares están muy próximos a la sangre de los capilares pulmonares.
El intercambio gaseoso entre el aire alveolar y la sangre pulmonar se produce a través de las membranas de todas las porciones terminales de los pulmones, no solo en los alvéolos. Todas estas membranas se conocen de manera colectiva como la membrana respiratoria.
Membrana respiratoria
Se pueden observar las siguientes capas de la membrana respiratoria:
Una capa de líquido que contiene surfactante y que tapiza el alvéolo, lo que reduce la tensión superficial del líquido alveolar.
El epitelio alveolar, que está formado por células epiteliales delgadas.
Una membrana basal epitelial.
Un espacio intersticial delgado entre el epitelio alveolar y la membrana capilar.
Una membrana basal capilar que en muchos casos se fusiona con la membrana basal del epitelio alveolar.
La membrana del endotelio capilar.
El grosor global de la membrana respiratoria en algunas zonas es tan pequeño como 0,2 μm, y en promedio es de aproximadamente 0,6 μm, excepto donde hay núcleos celulares se ha estimado que el área superficial total de la membrana respiratoria es de aproximadamente 70 m2 en el hombre adulto sano. La cantidad total de sangre en los capilares de los pulmones en cualquier instante dado es de 60 a 140 ml.
El diámetro medio de los capilares pulmonares es de solo aproximadamente 5 μm, lo que significa que los eritrocitos se deben comprimir a través de ellos. La membrana del eritrocito habitualmente toca la pared capilar, de modo que no es necesario que el O2 y el CO2 atraviesen cantidades significativas de plasma cuando difunden entre el alvéolo y el eritrocito. Esto también aumenta la rapidez de la difusión.
Factores que influyen en la velocidad de difusión gaseosa a través de la membrana respiratoria
Los factores que determinan la rapidez con la que un gas atraviesa la membrana son: 1) el grosor de la membrana; 2) el área superficial de la membrana; 3) el coeficiente de difusión del gas en la sustancia de la membrana, y 4) la diferencia de presión parcial del gas entre los dos lados de la membrana. De manera ocasional se produce un aumento del grosor de la membrana respiratoria, por ejemplo como consecuencia de la presencia de líquido de edema en el espacio intersticial de la membrana y en los alvéolos, de modo que los gases respiratorios deben difundir no solo a través de la membrana, sino también a través de este líquido.
El área superficial de la membrana respiratoria se puede reducir mucho en muchas situaciones. Por ejemplo, la resección de todo un pulmón reduce el área superficial total a la mitad de lo normal.
El coeficiente de difusión para la transferencia de cada uno de los gases a través de la membrana respiratoria depende de la solubilidad del gas en la membrana e inversamente de la raíz cuadrada del peso molecular del gas. La velocidad de difusión en la membrana respiratoria es casi exactamente la misma que en el agua, por los motivos que se han explicado antes. Por tanto, para una diferencia de presión dada, el CO2 difunde aproximadamente 20 veces más rápidamente que el O2. El oxígeno difunde aproximadamente dos veces más rápidamente que el nitrógeno. La diferencia de presión a través de la membrana respiratoria es la diferencia entre la presión parcial del gas en los alvéolos y la presión parcial del gas en la sangre capilar pulmonar. La presión parcial representa una medida del número total de moléculas de un gas particular que incide en una unidad de superficie de la superficie alveolar de la membrana por cada unidad de tiempo, y la presión del gas en la sangre representa el número de moléculas que intentarán escapar desde la sangre en la dirección opuesta. Por tanto, la diferencia entre estas dos presiones es una medida de la tendencia neta a que las moléculas del gas se muevan a través de la membrana. Cuando la presión parcial de un gas en los alvéolos es mayor que la presión del gas en la sangre, como ocurre en el caso del O2, se produce difusión neta desde los alvéolos hacia la sangre; cuando la presión del gas en la sangre es mayor que la presión parcial en los alvéolos, como ocurre en el caso del CO2, se produce difusión neta desde la sangre hacia los alvéolos.
Capacidad de difusión de la membrana respiratoria La capacidad de la membrana respiratoria de intercambiar un gas entre los alvéolos y la sangre pulmonar se expresa en términos cuantitativos por la capacidad de difusión de la membrana respiratoria, que se define como el volumen de un gas que difunde a través de la membrana en cada minuto para una diferencia de presión parcial de 1 mmHg. Todos los factores que se han analizado antes y que influyen en la difusión a través de la membrana respiratoria pueden influir sobre esta capacidad de difusión.
Capacidad de difusión del oxígeno En un hombre joven medio, la capacidad de difusión del O2 en condiciones de reposo es en promedio de 21 ml/min/mmHg. En términos funcionales.
La diferencia media de presión de O2 a través de la membrana respiratoria durante la respiración tranquila normal es de aproximadamente 11 mmHg. La multiplicación de esta presión por la capacidad de difusión (11 × 21) da un total de aproximadamente 230 ml de oxígeno que difunden a través de la membrana respiratoria cada minuto, que es igual a la velocidad a la que el cuerpo en reposo utiliza el O2. Aumento de la capacidad de difusión del oxígeno durante el ejercicio Durante el ejercicio muy intenso u otras situaciones que aumentan mucho el flujo sanguíneo pulmonar y la ventilación alveolar, la capacidad de difusión del O2 aumenta en los hombres jóvenes hasta un máximo de aproximadamente 65 ml/min/mmHg, que es el triple de la capacidad de difusión en situación de reposo.
Este aumento está producido por varios factores, entre los que se encuentran: 1) la apertura de muchos capilares pulmonares previamente cerrados o la dilatación adicional de capilares ya abiertos, aumentando de esta manera el área superficial de la sangre hacia la que puede difundir el O2.
2) un mejor equilibrio entre la ventilación de los alvéolos y la perfusión de los capilares alveolares con sangre, denominado cociente de ventilación-perfusión. Por tanto, durante el ejercicio la oxigenación de la sangre aumenta no solo por el aumento de la ventilación alveolar, sino también por una mayor capacidad de difusión de la membrana respiratoria para transportar el O2 hacia la sangre.
Capacidad de difusión del dióxido de carbono Nunca se ha medido la capacidad de difusión del CO2 porque el CO2 difunde a través de la membrana respiratoria con tanta rapidez que la Pco2 media de la sangre pulmonar no es muy diferente de la Pco2 de los alvéolos (la diferencia media es menor de 1 mmHg). Con las técnicas disponibles actualmente, esta diferencia es demasiado pequeña como para poderla medir. Sin embargo, las mediciones de la difusión de otros gases han mostrado que la capacidad de difusión varía directamente con el coeficiente de difusión del gas particular. Como el coeficiente de difusión del CO2 es algo mayor de 20 veces el del O2, cabe esperar que la capacidad de difusión del CO2 en reposo sea de aproximadamente 400 a 450 ml/min/mmHg y durante el esfuerzo de aproximadamente 1.200 a 1.300 ml/min/mmHg.
El pulmón tiene dos circulaciones: una circulación de bajo flujo y alta presión y una circulación de alto flujo y baja presión. La circulación de bajo flujo y alta presión aporta la sangre arterial sistémica a la tráquea, el árbol bronquial incluidos los bronquíolos terminales, los tejidos de sostén del pulmón y las capas exteriores (adventicias) de las arterias y venas pulmonares. Las arterias bronquiales, que son ramas de la aorta torácica, irrigan la mayoría de esta sangre arterial sistémica a una presión sólo ligeramente inferior a la presión aórtica. La circulación de alto flujo y baja presión que suministra la sangre venosa de todas las partes del organismo a los capilares alveolares en los que se añade el oxígeno (O2) y se extrae el dióxido de carbono (CO2). La arteria pulmonar, que recibe sangre del ventrículo derecho, y sus ramas arteriales transportan sangre a los capilares alveolares para el intercambio gaseoso y a las venas pulmonares y después devuelven la sangre a la aurícula izquierda para su bombeo por el ventrículo izquierdo a través de la circulación sistémica.
Anatomía fisiológica del sistema circulatorio pulmonar
Vasos pulmonares La arteria pulmonar se extiende solo 5 cm más allá de la punta del ventrículo derecho y después se divide en las ramas principales derecha e izquierda, que vascularizan los dos pulmones correspondientes. La arteria pulmonar tiene un grosor de pared un tercio del de la aorta. Las ramas de las arterias pulmonares son cortas, y todas las arterias pulmonares, incluso las arterias más pequeñas y las arteriolas, tienen diámetros mayores que sus correspondientes arterias sistémicas. Este aspecto, combinado con el hecho de que los vasos son delgados y distensibles, da al árbol arterial pulmonar una gran distensibilidad, que es en promedio de casi 7 ml/mmHg, que es similar a la de todo el árbol arterial sistémico. Esta gran distensibilidad permite que las arterias pulmonares se acomoden al gasto del volumen sistólico del ventrículo derecho. Las venas pulmonares, al igual que las arterias pulmonares, también son cortas. Drenan inmediatamente la sangre que les llega hacia la aurícula izquierda.
Vasos bronquiales La sangre también fluye hacia los pulmones a través de arterias bronquiales pequeñas que se originan en la circulación sistémica y transportan el 1-2% del gasto cardíaco total. Esta sangre arterial bronquial es sangre oxigenada, al contrario de la sangre parcialmente desoxigenada de las arterias pulmonares. Vascularizan los tejidos de soporte de los pulmones, como el tejido conjuntivo, los tabiques y los bronquios grandes y pequeños. Después de que esta sangre bronquial y arterial pase a través de los tejidos de soporte, drena hacia las venas pulmonares y entra en la aurícula izquierda, en lugar de regresar hacia la aurícula derecha. Por tanto, el flujo hacia la aurícula izquierda y el gasto del ventrículo izquierdo son aproximadamente un 1-2% mayores que el gasto del ventrículo derecho.
Linfáticos Hay vasos linfáticos en todos los tejidos de soporte del pulmón, comenzando en los espacios tisulares conjuntivos que rodean a los bronquíolos terminales, y siguiendo hacia el hilio del pulmón, y desde aquí principalmente hacia el conducto linfático torácico derecho. Las sustancias en forma de partículas que entran en los alvéolos son retiradas parcialmente por medio de estos conductos, y también eliminan de los tejidos pulmonares las proteínas plasmáticas que escapan de los capilares pulmonares, contribuyendo de esta manera a prevenir el edema pulmonar.
Presiones en el sistema pulmonar Estas curvas se comparan con la curva de presión aórtica, que es mucho más elevada. La presión sistólica del ventrículo derecho del ser humano normal es en promedio de aproximadamente 25 mmHg, y la presión diastólica es en promedio de aproximadamente 0 a 1 mmHg.
Presiones en la arteria pulmonar Durante la sístole la presión en la arteria pulmonar es esencialmente igual a la presión que hay en el ventrículo derecho. después del cierre de la válvula pulmonar al final de la sístole, la presión ventricular cae súbitamente, mientras que la presión arterial pulmonar disminuye más lentamente a medida que la sangre fluye a través de los capilares de los pulmones.
Presión capilar pulmonar Es de aproximadamente 7 mmHg. La importancia de esta baja presión capilar se analiza con más detalle más adelante en relación con las funciones de intercambio de líquidos de los capilares pulmonares.
Presiones auricular izquierda y venosa pulmonar La presión media en la aurícula izquierda y en las venas pulmonares principales es en promedio de aproximadamente 2 mmHg en el ser humano en decúbito, y varía desde un valor tan bajo como 1 mmHg hasta uno tan elevado como 5 mmHg.
La presión que se mide a través del catéter, denominada «presión de enclavamiento», es de aproximadamente 5 mmHg. Como todo el flujo sanguíneo se ha interrumpido en la arteria pequeña enclavada, y como los vasos sanguíneos que se extienden más allá de esta arteria establecen una conexión directa con los capilares pulmonares, esta presión de enclavamiento es habitualmente solo de 2 a 3 mmHg mayor que la presión auricular izquierda. Cuando la presión auricular izquierda aumenta a valores elevados, también lo hace la presión de enclavamiento pulmonar. Por tanto, las mediciones de la presión de enclavamiento se pueden utilizar para estudiar las alteraciones de la presión capilar pulmonar y de la presión auricular izquierda en pacientes que tienen insuficiencia cardíaca congestiva.
Volumen sanguíneo de los pulmones El volumen de la sangre de los pulmones es de aproximadamente 450 ml, aproximadamente el 9% del volumen de sangre total de todo el aparato circulatorio. Aproximadamente 70 ml de este volumen de sangre pulmonar están en los capilares pulmonares, y el resto se divide aproximadamente por igual entre las arterias y las venas pulmonares.
Los pulmones sirven como reservorio de sangre En varias situaciones fisiológicas y patológicas la cantidad de sangre de los pulmones puede variar desde tan poco como la mitad del valor normal hasta el doble de lo normal. Por ejemplo, cuando una persona sopla aire con tanta intensidad que se genera una presión elevada en los pulmones (como cuando se toca una trompeta), se pueden expulsar hasta 250 ml de sangre desde el aparato circulatorio pulmonar hacia la circulación sistémica. Por otro lado, la pérdida de sangre desde la circulación sistémica por una hemorragia puede ser compensada parcialmente por el desplazamiento automático de sangre desde los pulmones hacia los vasos sistémicos. La patología cardíaca puede desplazar sangre desde la circulación sistémica a la circulación pulmonar La insuficiencia del lado izquierdo del corazón o el aumento de la resistencia al flujo sanguíneo a través de la válvula mitral como consecuencia de una estenosis mitral o una insuficiencia mitral hace que la sangre quede estancada en la circulación pulmonar, aumentando a veces el volumen de sangre pulmonar hasta un 100% y produciendo grandes aumentos de las presiones vasculares pulmonares. Dado que el volumen de la circulación sistémica es aproximadamente nueve veces el de la circulación pulmonar, el desplazamiento de sangre desde un sistema hacia el otro afecta mucho al sistema pulmonar, pero habitualmente tiene solo efectos circulatorios sistémicos leves.
Zonas 1, 2 y 3 del flujo sanguíneo pulmonar Los capilares de las paredes alveolares están distendidos por la presión de la sangre que hay en su interior, pero simultáneamente están comprimidos por la presión del aire alveolar que está en su exterior. Por tanto, siempre que la presión del aire alveolar pulmonar sea mayor que la presión de la sangre capilar, los capilares se cierran y no hay flujo sanguíneo. En diferentes situaciones normales y patológicas se puede encontrar una cualquiera de tres posibles zonas (patrones) del flujo sanguíneo pulmonar, como se señala a continuación: Zona 1: ausencia de flujo durante todas las porciones del ciclo cardíaco porque la presión capilar alveolar local en esa zona del pulmón nunca aumenta por encima de la presión del aire alveolar en ninguna fase del ciclo cardíaco. Zona 2: flujo sanguíneo intermitente, solo durante los picos de presión arterial pulmonar, porque la presión sistólica en ese momento es mayor que la presión del aire alveolar, pero la presión diastólica es menor que la presión del aire alveolar. Zona 3: flujo de sangre continuo, porque la presión capilar alveolar es mayor que la presión del aire alveolar durante todo el ciclo cardíaco. Normalmente los pulmones solo tienen flujo sanguíneo en las zonas 2 y 3, la zona 2 (flujo intermitente) en los vértices y la zona 3 (flujo continuo) en todas las zonas inferiores. Por ejemplo, cuando una persona está en posición erguida la presión arterial pulmonar en el vértice pulmonar es aproximadamente 15 mmHg menor que la presión a nivel del corazón. Por tanto, la presión sistólica apical es de solo 10 mmHg (25 mmHg a nivel del corazón menos la diferencia de presión hidrostática de 15 mmHg). Esta presión sanguínea apical de 10 mmHg es mayor que la presión cero del aire alveolar, de modo que la sangre fluye a través de los capilares apicales pulmonares durante la sístole cardíaca. Por el contrario, durante la diástole la presión diastólica de 8 mmHg a nivel del corazón no es suficiente para empujar la sangre contra el gradiente de presión hidrostática de 15 mmHg necesario para producir el flujo capilar diastólico. Por tanto, el flujo sanguíneo a través de la parte apical del pulmón es intermitente, de modo que hay flujo durante la sístole e interrupción del flujo durante la diástole; esto se denomina flujo sanguíneo de zona 2. El flujo sanguíneo de zona 2 comienza en los pulmones normales aproximadamente 10 cm por encima del nivel medio del corazón y se extiende desde ahí hasta la parte superior de los pulmones. En las regiones inferiores de los pulmones, desde aproximadamente 10 cm por encima del nivel del corazón hasta la parte inferior de los pulmones, la presión arterial pulmonar durante la sístole y la diástole es mayor que la presión del aire alveolar, que es cero. Por tanto, existe un flujo continuo a través de los capilares pulmonares alveolares, o flujo sanguíneo de zona 3. Además, cuando una persona está tumbada, no hay ninguna parte del pulmón que esté más de algunos centímetros por encima del nivel del corazón. En este caso el flujo sanguíneo de una persona normal es totalmente un flujo sanguíneo de zona 3, incluyendo los vértices pulmonares.
El aumento del gasto cardíaco durante el ejercicio intenso es asumido normalmente por la circulación pulmonarsin grandes aumentos en la presión arterial pulmonar Durante el ejercicio intenso el flujo sanguíneo a través de los pulmones puede aumentar entre cuatro y siete veces. Este flujo adicional se acomoda en los pulmones de tres formas:
1) aumentando el número de capilares abiertos, a veces hasta tres veces
2) distendiendo todos los capilares y aumentando la velocidad del flujo a través de cada capilar a más del doble
3) aumentando la presión arterial pulmonar. Normalmente, las dos primeras modificaciones reducen la resistencia vascular pulmonar tanto que la presión arterial pulmonar aumenta muy poco, incluso durante el ejercicio máximo.
Dinámica capilar pulmonar Las paredes alveolares están tapizadas por tantos capilares que en la mayor parte de los sitios los capilares casi se tocan entre sí, adosados unos a otros. Por tanto, con frecuencia se dice que la sangre capilar fluye en las paredes alveolares como una «lámina de flujo», y no como capilares individuales.
Presión capilar pulmonar Nunca se han realizado mediciones directas de la presión capilar pulmonar. Sin embargo, el método «isogravimétrico» de la presión capilar pulmonar, ha dado un valor de 7 mmHg. Es probable que esta medida sea casi correcta, porque la presión auricular izquierda media es de aproximadamente 2mmHg y la presión arterial pulmonar media es de solo 15 mmHg, de modo que la presión capilar pulmonar media debe estar en algún punto entre estos dos valores.
Duración del tiempo que la sangre permanece en los capilares pulmonares A partir del estudio histológico del área transversal total de todos los capilares pulmonares se puede calcular que cuando el gasto cardíaco es normal la sangre pasa a través de los capilares pulmonares en aproximadamente 0,8 s. Cuando aumenta el gasto cardíaco este tiempo puede acortarse hasta 0,3 s. Este acortamiento sería mucho mayor si no fuera por el hecho de que se abren capilares adicionales, que normalmente están colapsados, para acomodarse al aumento del flujo sanguíneo. Así, en solo una fracción de segundo la sangre que pasa a través de los capilares alveolares se oxigena y pierde su exceso de dióxido de carbono.
Líquido en la cavidad pleural Cuando los pulmones se expanden y se contraen durante la respiración normal se deslizan en el interior de la cavidad pleural. Para facilitar este movimiento hay una delgada capa de líquido mucoide entre las pleuras parietal y visceral.
«Presión negativa» en el líquido pleural Siempre es necesaria una fuerza negativa en el exterior de los pulmones para mantener expandidos los pulmones. Esta fuerza es proporcionada por la presión negativa del espacio pleural normal. La causa básica de esta presión negativa es el bombeo de líquidos desde el espacio pleural por los linfáticos (que también es la base de la presión negativa que se encuentra en la mayor parte de los espacios tisulares del cuerpo). Como la tendencia al colapso normal de los pulmones es de aproximadamente –4 mmHg, la presión del líquido pleural siempre debe ser al menos tan negativa como –4 mmHg para mantener expandidos los pulmones. Las mediciones reales han mostrado que la presión habitualmente es de aproximadamente –7 mmHg, que es algunos mmHg más negativa que la presión de colapso de los pulmones. Así, la negatividad de la presión del líquido pleural mantiene los pulmones normales traccionados contra la pleura parietal de la cavidad torácica, excepto por una capa muy delgada de líquido mucoide que actúa como lubricante.
Derrame pleural: acumulación de grandes cantidades de líquido libre en el espacio pleural El derrame es análogo al líquido de edema en los tejidos y se puede denominar «edema de la cavidad pleural». Las causas del derrame son las mismas que las causas del edema en otros tejidos entre ellas:
1) bloqueo del drenaje linfático desde la cavidad pleural.
2)insuficiencia cardíaca, que da lugar a unas presiones capilares periférica y pulmonar excesivamente altas, que dan lugar a una trasudación excesiva de líquido hacia la cavidad pleural.
3) marcada reducción de la presión osmótica coloidal del plasma, que permite una trasudación excesiva de líquidos.
4) infección o cualquier otra causa de inflamación de las superficies de la cavidad pleural, que aumenta la permeabilidad de las membranas capilares y permite la salida rápida tanto de proteínas plasmáticas como de líquido hacia la cavidad.
Mecánica de la ventilación pulmonar Músculos que causan la expansión y contracción pulmonar Los pulmones se pueden expandir y contraer de dos maneras:
1) mediante el movimiento hacia abajo y hacia arriba del diafragma para alargar o acortar la cavidad torácica,
2) mediante la elevación y el descenso de las costillas para aumentar y reducir el diámetro anteroposterior de la cavidad torácica.
La respiración tranquila normal se consigue casi totalmente por el primer mecanismo, es decir, por el movimiento del diafragma. Durante la inspiración la contracción del diafragma tira hacia abajo de las superficies inferiores de los pulmones. Después, durante la espiración el diafragma simplemente se relaja, y el retroceso elástico de los pulmones, de la pared torácica y de las estructuras abdominales comprime los pulmones y expulsa el aire. Sin embargo, durante la respiración forzada las fuerzas elásticas no son suficientemente potentes para producir la espiración rápida necesaria, de modo que se consigue una fuerza adicional principalmente mediante la contracción de los músculos abdominales, que empujan el contenido abdominal hacia arriba contra la parte inferior del diafragma, comprimiendo de esta manera los pulmones.
El segundo método para expandir los pulmones es elevar la caja torácica. Al elevarla se expanden los pulmones porque, en la posición de reposo natural, las costillas están inclinadas hacia abajo.
Los músculos más importantes que elevan la caja torácica son los intercostales externos, aunque otros músculos que contribuyen son:
1) los músculos esternocleidomastoideos, que elevan el esternón
2) los serratos anteriores, que elevan muchas de las costillas
3) los escalenos, que elevan las dos primeras costillas. Los músculos que tiran hacia abajo de la caja costal durante la espiración son principalmente 1) los rectos del abdomen, que tienen el potente efecto de empujar hacia abajo las costillas inferiores al mismo tiempo que ellos y otros músculos abdominales también comprimen el contenido abdominal hacia arriba contra el diafragma, y 2) los intercostales internos.
ESTERNOCLEIDOMASTOIDEOSERRATOSESCALENO
Presiones que originan el movimiento de entrada y salida de aire de los pulmones El pulmón es una estructura elástica que se colapsa como un globo y expulsa el aire a través de la tráquea siempre que no haya ninguna fuerza que lo mantenga insuflado. Además, no hay uniones entre el pulmón y las paredes de la caja torácica, excepto en el punto en el que está suspendido del mediastino, la sección media de la cavidad torácica, en el hilio. Por el contrario, el pulmón «flota» en la cavidad torácica, rodeado por una capa delgada de líquido pleural que lubrica el movimiento de los pulmones en el interior de la cavidad. Además, la aspiración continua del exceso de líquido hacia los conductos linfáticos mantiene una ligera presión negativa entre la superficie visceral del pulmón y la superficie pleural parietal de la cavidad torácica. Por tanto, los pulmones están sujetos a la pared torácica como si estuvieran pegados, excepto porque están bien lubricados y se pueden deslizar libremente cuando el tórax se expande y se contrae.
Presión pleural y sus cambios durante la respiración La presión pleural es la presión del líquido que está en el delgado espacio que hay entre la pleura pulmonar y la pleura de la pared torácica. Como se ha señalado antes, esta presión es normalmente una aspiración ligera, lo que significa que hay una presión ligeramente negativa. La presión pleural normal al comienzo de la inspiración es de aproximadamente –5 cmH2O, que es la magnitud de la aspiración necesaria para mantener los pulmones expandidos hasta su nivel de reposo. Durante la inspiración normal, la expansión de la caja torácica tira hacia fuera de los pulmones con más fuerza y genera una presión más negativa, hasta un promedio de aproximadamente –7,5 cmH2O.
Presión alveolar: presión del aire en el interior de los alvéolos pulmonares Cuando la glotis está abierta y no hay flujo de aire hacia el interior ni el exterior de los pulmones, las presiones en todas las partes del árbol respiratorio, hasta los alvéolos, son iguales a la presión atmosférica, que se considera que es la presión de referencia cero en las vías aéreas (es decir, presión de 0 cmH2O). Para que se produzca un movimiento de entrada de aire hacia los alvéolos durante la inspiración, la presión en los alvéolos debe disminuir hasta un valor ligeramente inferior a la presión atmosférica (debajo de cero). La segunda curva (denominada «presión alveolar») muestra que durante la inspiración normal la presión alveolar disminuye hasta aproximadamente –1 cmH2O. Esta ligera presión negativa es suficiente para arrastrar 0,5 l de aire hacia los pulmones en los 2 s necesarios para una inspiración tranquila normal. Durante la espiración, la presión alveolar aumenta hasta aproximadamente +1 cmH2O, lo que fuerza la salida del 0,5 l de aire inspirado desde los pulmones durante los 2 a 3 s de la espiración.
Presión transpulmonar: diferencia entre las presiones alveolar y pleural Se observa que la presión transpulmonar es la diferencia entre la presión que hay en el interior de los alvéolos y la que hay en las superficies externas de los pulmones (presión pleural), y es una medida de las fuerzas elásticas de los pulmones que tienden a colapsarlos en todos los momentos de la respiración, denominadas presión de retroceso.
Distensibilidad de los pulmones El volumen que se expanden los pulmones por cada aumento unitario de presión transpulmonar (si se da tiempo suficiente para alcanzar el equilibrio) se denomina distensibilidad pulmonar. La distensibilidad pulmonar total de los dos pulmones en conjunto en el ser humano adulto normal es en promedio de aproximadamente 200 ml de aire por cada cmH2O de presión transpulmonar. Es decir, cada vez que la presión transpulmonar aumenta 1 cmH2O, el volumen pulmonar, después de 10 a 20 s, se expande 200 ml.
Surfactante, tensión superficial y colapso de los alvéolos
Principio de la tensión superficial Cuando el agua forma una superficie con el aire, las moléculas de agua de la superficie del agua tienen una atracción especialmente intensa entre sí. En consecuencia, la superficie del agua siempre está intentando contraerse. Esto es lo que mantiene unidas entre sí las gotas de lluvia: una membrana muy contráctil de moléculas de agua que rodea toda la superficie de la gota de agua. Invirtamos ahora estos principios y veamos qué ocurre en las superficies internas de los alvéolos. Aquí la superficie de agua también intenta contraerse, lo que tiende a expulsar el aire de los alvéolos a través de los bronquios y, al hacerlo, hace que los alvéolos intentan colapsar. El efecto neto es producir una fuerza contráctil elástica de todo el pulmón, que se denomina fuerza elástica de la tensión superficial.
El surfactante y su efecto sobre la tensión superficial El surfactante es un agente activo de superficie en agua, lo que significa que reduce mucho la tensión superficial del agua. Es secretado por células epiteliales especiales secretoras de surfactante denominadas células epiteliales alveolares de tipo II, que constituyen aproximadamente el 10% del área superficial de los alvéolos. Estas células son granulares y contienen inclusiones de lípidos que se secretan en el surfactante hacia los alvéolos. El surfactante es una mezcla compleja de varios fosfolípidos, proteínas e iones.
Efecto de la caja torácica sobre la expansibilidad pulmonar Hasta ahora hemos analizado la capacidad de expansión de los pulmones de manera aislada, sin considerar la caja torácica. La caja torácica tiene sus propias características elásticas y viscosas, similares a las de los pulmones; incluso si los pulmones no estuvieran presentes en el tórax, seguiría siendo necesario un esfuerzo muscular para expandir la caja torácica. Distensibilidad del tórax y de los pulmones en conjunto
La distensibilidad de todo el sistema pulmonar
(los pulmones y la caja torácica en conjunto) se mide cuando se expanden los pulmones de una persona relajada o paralizada totalmente. Para medir la distensibilidad se introduce aire en los pulmones poco a poco mientras se registran las presiones y volúmenes pulmonares. Para insuflar este sistema pulmonar total es necesario casi el doble de presión que para insuflar los mismos pulmones después de extraerlos de la caja torácica. Por tanto, la distensibilidad del sistema pulmón-tórax combinado es casi exactamente la mitad que la de los pulmones solos, 110 ml de volumen por cada cmH2O de presión para el sistema combinado, en comparación con 200 ml/cmH2O para los pulmones de manera aislada.
Volúmenes y capacidades pulmonares
Registro de las variaciones del volumen pulmonar: espirometría La ventilación pulmonar puede estudiarse registrando el movimiento del volumen del aire que entra y sale de los pulmones, un método que se denomina espirometría.
VOLÚMENES PULMONARES
El volumen corriente es el volumen de aire que se inspira o se espira en cada respiración normal; es igual a aproximadamente 500 ml en el hombre adulto medio.
El volumen de reserva inspiratoria es el volumen adicional de aire que se puede inspirar desde un volumen corriente normal y por encima del mismo cuando la persona inspira con una fuerza plena; habitualmente es igual a aproximadamente 3.000 ml.
El volumen de reserva espiratoria es el volumen adicional máximo de aire que se puede espirar mediante una espiración forzada después del final de una espiración a volumen corriente normal; normalmente, este volumen es igual a aproximadamente 1.100 ml.
El volumen residual es el volumen de aire que queda en los pulmones después de la espiración más forzada; este volumen es en promedio de aproximadamente 1.200 ml
Determinación de la capacidad residual funcional, el volumen residual y la capacidad pulmonar total: método de dilución de helio
La capacidad residual funcional (CRF), que es el volumen de aire que queda en los pulmones al final de una espiración normal, es importante en la función pulmonar. Como su valor se altera mucho en algunos tipos de enfermedad pulmonar, con frecuencia es deseable medir esta capacidad.
Se llena un espirómetro de volumen conocido con aire mezclado con helio a una concentración conocida. Antes de respirar del espirómetro la persona hace una espiración normal. Al final de esta espiración, el volumen que queda en los pulmones es igual a la CRF. En este punto el paciente comienza inmediatamente a respirar desde el espirómetro, y los gases del espirómetro se mezclan con los gases de los pulmones. En consecuencia, el helio es diluido por los gases de la CRF, y se puede calcular el volumen de la CRF a partir del grado de dilución del helio.
El volumen respiratorio minuto equivale a la frecuencia respiratoria multiplicada por el volumen corriente El volumen respiratorio minuto es la cantidad total de aire nuevo que pasa hacia las vías aéreas en cada minuto y es igual al volumen corriente multiplicado por la frecuencia respiratoria por minuto. El volumen corriente normal es de aproximadamente 500 ml y la frecuencia respiratoria normal es de aproximadamente 12 respiraciones por minuto. Por tanto, el volumen respiratorio minuto es en promedio de aproximadamente 6 l/min. Una persona puede vivir durante un período breve con un volumen respiratorio minuto de tan solo 1,5 l/min y una frecuencia respiratoria de solo 2 a 4 respiraciones por minuto. La frecuencia respiratoria aumenta de manera ocasional a 40 a 50 por minuto, y el volumen corriente se puede hacer tan grande como la capacidad vital, aproximadamente 4.600 ml en un hombre adulto joven. Esto puede dar un volumen respiratorio minuto mayor de 200 l/min, o más de 30 veces el valor normal. La mayor parte de las personas no puede mantener más de la mitad a dos tercios de estos valores durante más de 1 min.
Ventilación alveolar En último término, la función de la ventilación pulmonar es renovar continuamente el aire de las zonas de intercambio gaseoso de los pulmones, en las que el aire está próximo a la sangre pulmonar. Estas zonas incluyen los alvéolos, los sacos alveolares, los conductos alveolares y los bronquiolos respiratorios. La velocidad a la que llega a estas zonas el aire nuevo se denomina ventilación alveolar.

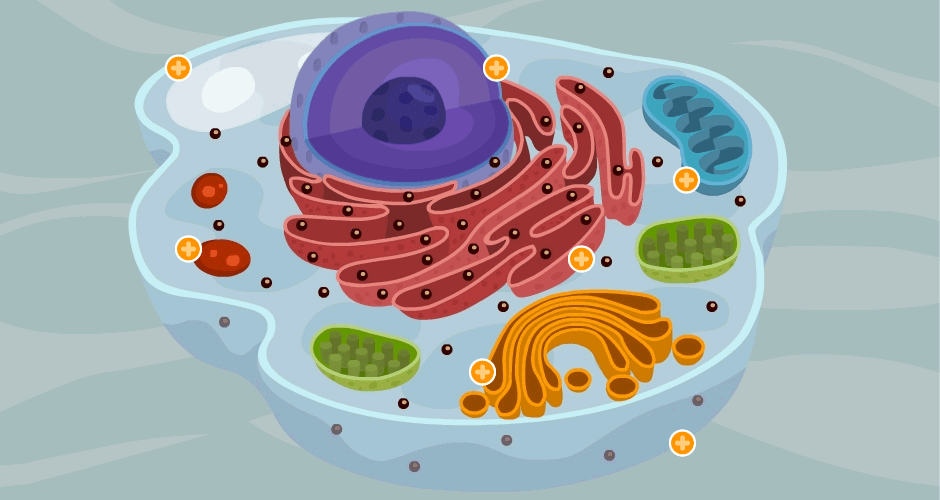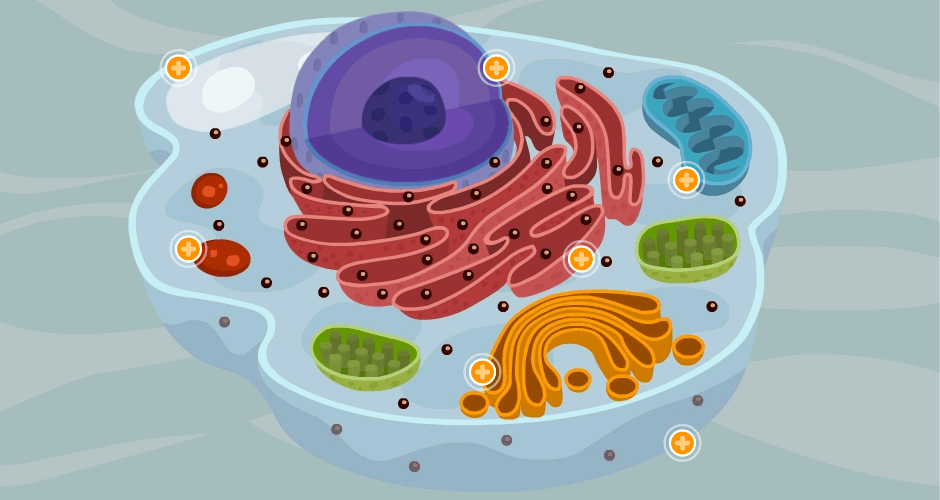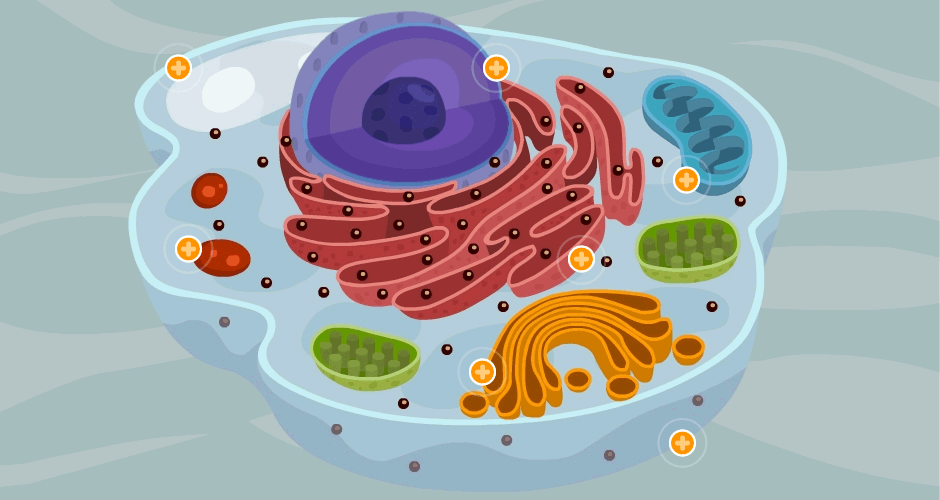
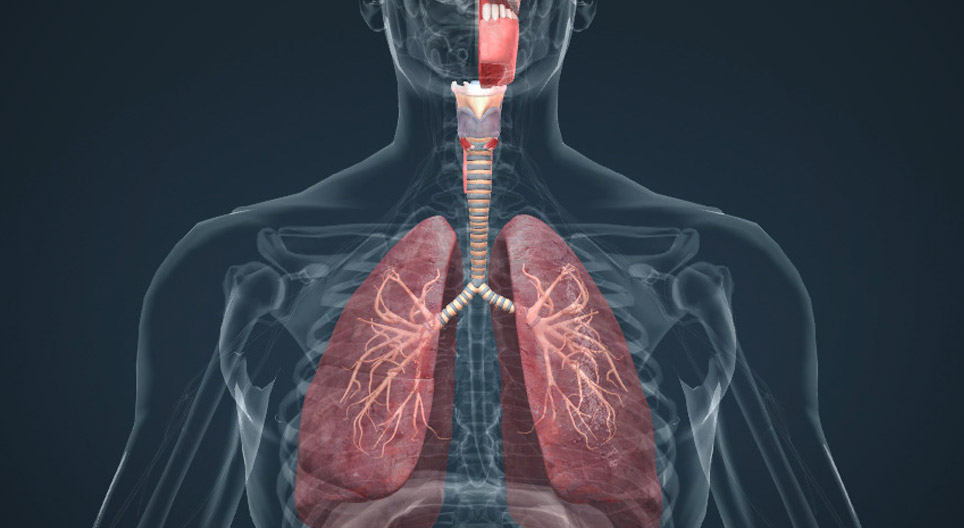

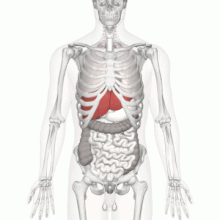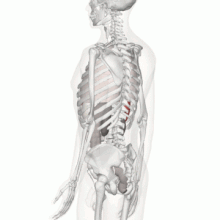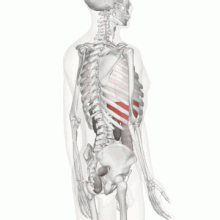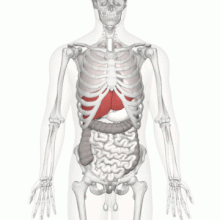




















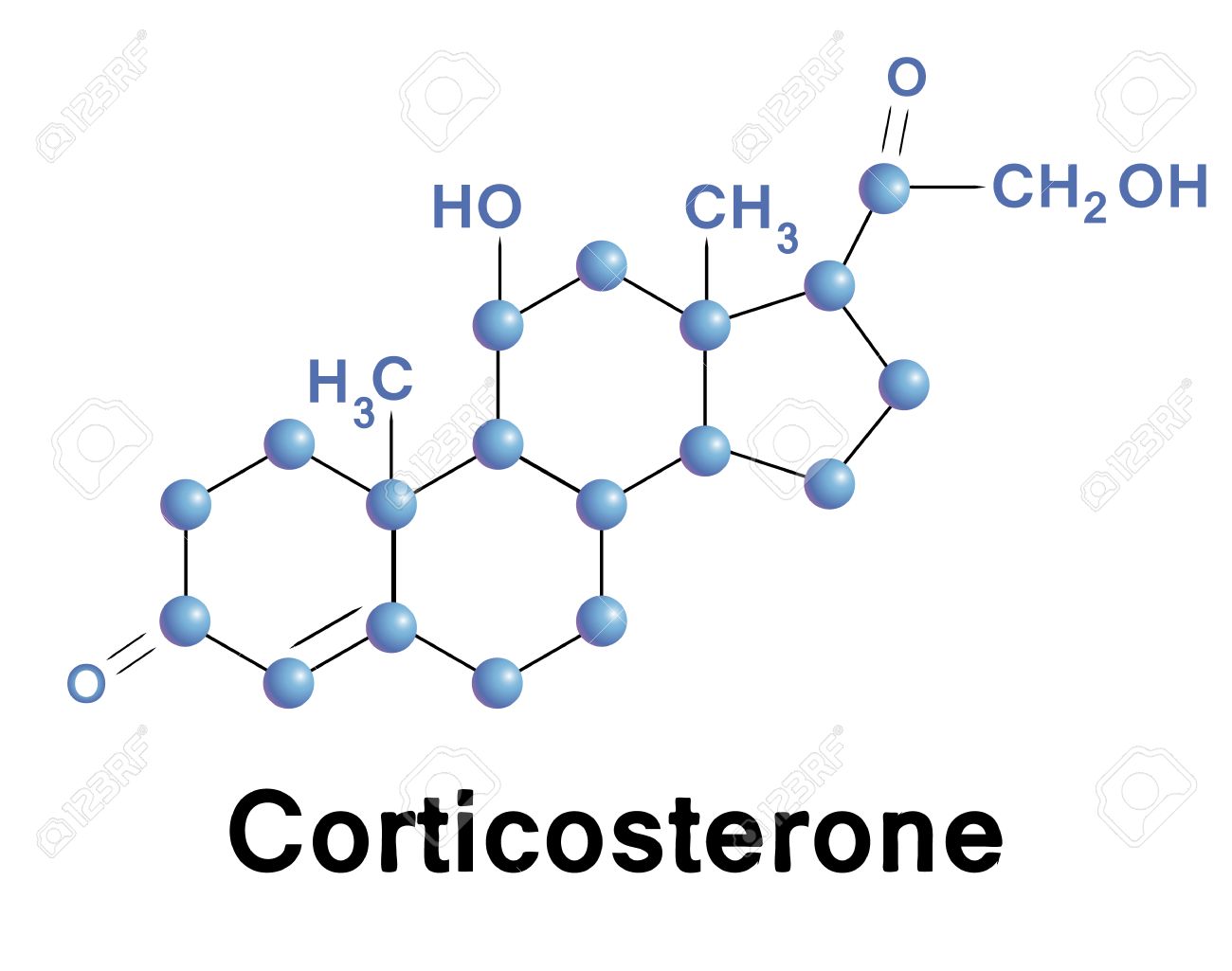



![Qué es ACTH? » Su Definición y Significado [2020]](https://conceptodefinicion.de/wp-content/uploads/2014/04/acth.jpg)




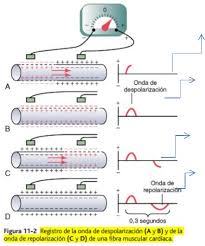
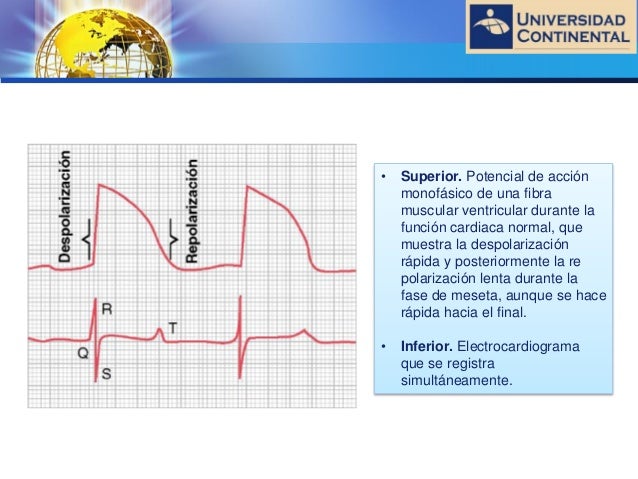


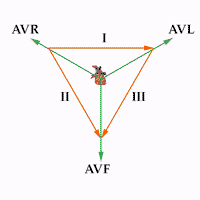
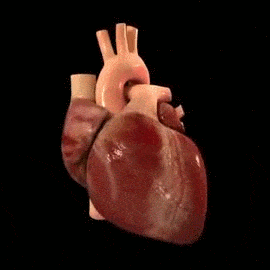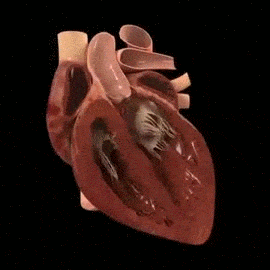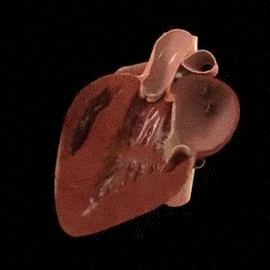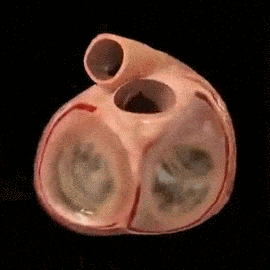


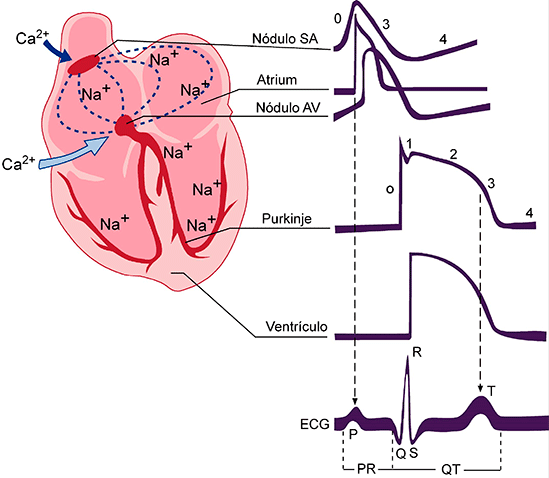



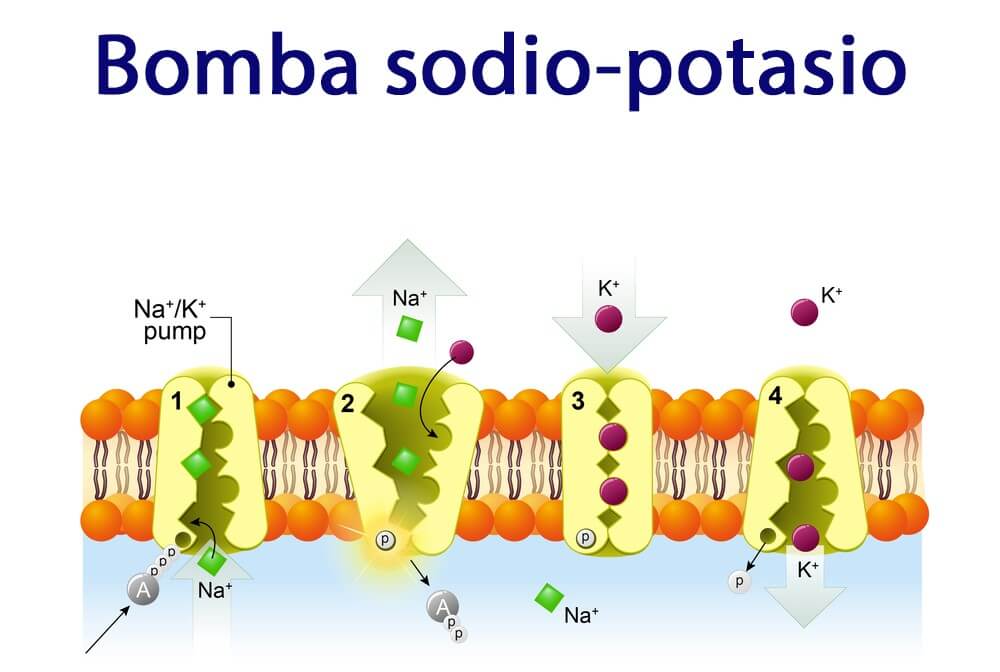
/como-es-el-corazon-597b60145f9b58928bd8c808.jpg)