Corteza cerebral, funciones intelectuales del cerebro, aprendizaje y memoria.
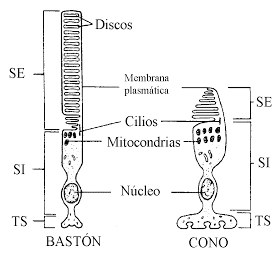
El elemento funcional de la corteza cerebral es una fina capa de neuronas que cubre la superficie de todas las circunvoluciones del cerebro. Esta capa solo tiene un grosor de 2 a 5 mm, y el área total que ocupa mide más o menos la cuarta parte de un metro cuadrado. En total, la corteza cerebral contiene unos 100.000 millones de neuronas. La estructura histológica típica de la superficie neuronal de la corteza cerebral, con sus sucesivas capas formadas por diversos tipos de neuronas. La mayor parte de estas células son de tres tipos:
1) células de los granos (que también se denominan células estrelladas)
2) fusiformes
3) piramidales, las cuales reciben su nombre por su característica forma piramidal.
Las células de los granos en general tienen axones cortos y, por tanto, funcionan básicamente como interneuronas que nada más transmiten señales nerviosas hasta una distancia corta en el interior de la corteza. Algunas son excitadoras y liberan sobre todo el neurotransmisor excitador glutamato, mientras que otras son inhibidoras y dejan salir especialmente el neurotransmisor inhibidor ácido γ-aminobutírico (GABA).

Las áreas sensitivas de la corteza así como las áreas de asociación entre ellas y las motoras poseen grandes concentraciones de estas células de los granos, lo que quiere decir que existe un alto grado de procesamiento intracortical de las señales sensitivas recibidas en el seno de las áreas sensitivas y de asociación. Las células piramidales y fusiformes dan lugar a casi todas las fibras de salida desde la corteza. Las piramidales, que tienen un mayor tamaño y son más abundantes que las fusiformes, constituyen la fuente de las fibras nerviosas grandes y largas que recorren toda la médula espinal. Las células piramidales también originan la mayoría de los amplios haces de fibras de asociación subcorticales que van desde una parte principal del encéfalo a otra.
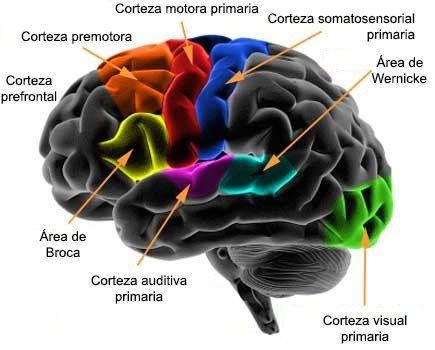
Funciones cumplidas por áreas corticales específicas.
La reunión de grandes cantidades de información procedentes de muchas fuentes distintas produce un mapa más general. Las áreas motoras primarias poseen conexiones directas con músculos específicos para originar movimientos musculares concretos. Las áreas sensitivas primarias detectan sensaciones concretas (visual, auditiva o somática) que se transmiten directamente hasta el cerebro desde los órganos sensitivos periféricos.
Las áreas secundarias interpretan las señales procedentes de las áreas primarias. Por ejemplo, las áreas premotora y suplementaria funcionan junto con la corteza motora primaria y los ganglios basales para suministrar «patrones» de actividad motora. En el ámbito de los sentidos, las áreas sensitivas secundarias, situadas a unos centímetros de distancia de las primarias, comienzan a analizar los significados de las señales sensitivas concretas, como por ejemplo: 1) la interpretación de la forma y la textura de un objeto cogido con la mano; 2) la interpretación del color, la intensidad lumínica, las direcciones de las líneas y los ángulos y otros aspectos de la visión, y 3) la interpretación de los significados que tienen los tonos sonoros y sus secuencias en las señales auditivas.
Área de asociación parietooccipitotemporal.
El área de asociación parietooccipitotemporal está situada en el gran espacio de la corteza parietal y occipital cuyo límite anterior corresponde a la corteza somatosensitiva, el posterior a la corteza visual y el lateral a la corteza auditiva. Según cabría esperar, proporciona un alto grado de significación interpretativa a las señales procedentes de todas las áreas sensitivas que la rodean. Sin embargo, hasta el área de asociación parietooccipitotemporal posee sus propias subáreas funcionales.
El área de Wernicke es importante para la comprensión del lenguaje.
El área principal para la comprensión del lenguaje, denominada área de Wernicke, está detrás de la corteza auditiva primaria en la parte posterior de la circunvolución superior del lóbulo temporal. Más adelante la explicamos con más detalle; se trata de la región más importante de todo el cerebro para las funciones intelectuales superiores porque casi todas ellas están basadas en el lenguaje.
Área de asociación prefrontal
El área de asociación prefrontal funciona en íntima asociación con la corteza motora para planificar los patrones complejos y las secuencias de los actos motores. Como contribución a esta actividad, recibe potentes señales aferentes a través de un enorme haz subcortical de fibras nerviosas que conectan el área de asociación parietooccipitotemporal con el área de asociación prefrontal. Por esta vía, la corteza prefrontal recibe mucha información sensitiva sometida ya a un primer análisis, referida especialmente a las coordenadas espaciales del cuerpo, que hace falta para planificar unos movimientos eficaces. Gran parte de los impulsos emitidos desde el área prefrontal hacia el sistema de control motor atraviesan la porción correspondiente al caudado dentro del circuito de retroalimentación para la planificación motora establecido entre los ganglios basales y el tálamo, lo que aporta muchos de los ingredientes secuenciales y paralelos para la estimulación del movimiento.

Área de asociación límbica.
Está situada en el polo anterior del lóbulo temporal, en la porción ventral del lóbulo frontal y en la circunvolución cingular que queda en la profundidad de la cisura longitudinal por la cara medial de cada hemisferio cerebral. Se ocupa sobre todo del comportamiento, las emociones y la motivación. Este sistema límbico proporciona la mayoría de los impulsos emocionales para activar otras áreas del encéfalo e incluso suministra el estímulo encargado de motivar el propio proceso de aprendizaje.
Área para el reconocimiento de las caras.
Un tipo de alteración cerebral interesante llamada prosopagnosia consiste en la incapacidad para reconocer las caras. Este trastorno sucede en personas con una amplia lesión en la parte inferomedial de ambos lóbulos occipitales además de en las caras medioventrales de los lóbulos temporales. La pérdida de estas áreas destinadas al reconocimiento facial, aunque parezca mentira, propicia pocas alteraciones más del funcionamiento cerebral.
Concepto de hemisferio dominante.
Las funciones interpretativas generales del área de Wernicke y de la circunvolución angular, así como las funciones que cumplen las áreas del lenguaje y de control motor, suelen estar mucho más desarrolladas en un hemisferio cerebral que en el otro; por consiguiente, en este lado recibe el nombre de hemisferio dominante. Más o menos en el 95% de las personas, el hemisferio dominante es el izquierdo. Desde el momento del parto, la superficie cortical que con el tiempo acabará convirtiéndose en el área de Wernicke llega a ser un 50% mayor en el hemisferio izquierdo que en el derecho en más de la mitad de los recién nacidos. Por tanto, no cuesta entender por qué el lado izquierdo del cerebro podría volverse dominante sobre el derecho. Sin embargo, si por alguna razón esta área queda dañada o anulada muy al principio de la infancia, el lado opuesto del cerebro normalmente adquirirá la posición dominante.
Funciones de la corteza parietooccipitotemporal en el hemisferio no dominante.
Cuando el área de Wernicke queda destruida en el hemisferio dominante de un adulto, la persona normalmente pierde casi todas las funciones intelectuales asociadas al lenguaje o al simbolismo verbal, como la capacidad para leer, para efectuar operaciones matemáticas, e incluso para pensar en el caso de los problemas lógicos. En cambio, se conservan otros muchos tipos de capacidades interpretativas, algunas de las cuales recurren a las regiones del lóbulo temporal y la circunvolución angular del hemisferio opuesto.
Pensamientos, conciencia y memoria.
Nuestro problema más difícil al abordar la conciencia, el pensamiento, la memoria y el aprendizaje radica en que ignoramos los mecanismos nerviosos que sigue un pensamiento y conocemos poco sobre el mecanismo de la memoria. Sí que sabemos que la destrucción de grandes porciones de la corteza cerebral no impide que una persona tenga pensamientos, pero sí reduce su profundidad y también el grado de conciencia que ejerce sobre su medio. No hay duda de que cada pensamiento entraña unas señales simultáneas en muchas porciones de la corteza cerebral, el tálamo, el sistema límbico y la formación reticular del tronco del encéfalo.
Algunos pensamientos básicos probablemente dependan casi por completo de los centros inferiores; la idea del dolor quizá sea un buen ejemplo debido a que la estimulación eléctrica de la corteza humana rara vez suscita algo más que un dolor leve, mientras que en el caso de ciertas regiones del hipotálamo, la amígdala y el mesencéfalo puede provocar un dolor atroz. Por el contrario, un tipo de patrón de pensamiento que requiere una gran participación de la corteza cerebral es el de la visión, debido a que la ausencia de la corteza visual genera una absoluta incapacidad para percibir las formas visuales o los colores.
Memoria: funciones de la facilitación y la inhibición sinápticas.
Los recuerdos se almacenan en el cerebro al variar la sensibilidad básica de la transmisión sináptica entre las neuronas como consecuencia de la actividad nerviosa previa. Las vías nuevas o facilitadas se llaman huellas de memoria. Son importantes porque, una vez que quedan establecidas, es posible activarlas de forma selectiva por los pensamientos de la mente para reproducir los recuerdos. Los experimentos con los animales inferiores han puesto de manifiesto que las huellas de memoria pueden darse a cualquier nivel del sistema nervioso. Hasta los reflejos medulares varían al menos un poco como respuesta a la activación repetida de la médula, y estos cambios reflejos forman parte del proceso de la memoria. Asimismo, los recuerdos a largo plazo derivan de modificaciones producidas en la conducción sináptica de los centros cerebrales inferiores. Sin embargo, la mayor parte de los recuerdos que asociamos a los mecanismos intelectuales se basan en las huellas de memoria de la corteza cerebral.
Memoria a corto plazo.
La memoria a corto plazo viene representada por el recuerdo de las 7 a 10 cifras que forman un número de teléfono (o de 7 a 10 hechos independientes diferentes) durante unos pocos segundos o minutos en un momento dado, pero que solo dura mientras la persona siga pensando en dichos números o en dichas circunstancias.
Memoria a medio plazo.
Los recuerdos a medio plazo pueden durar muchos minutos o incluso semanas. A la larga desaparecerán a no ser que se activen suficientes huellas de memoria como para volverse más permanentes; en ese momento, se clasificarán como recuerdos a largo plazo. Los experimentos con animales primitivos han demostrado que el tipo de los recuerdos a medio plazo puede obedecer a cambios físicos o químicos transitorios o a ambos procesos, ocurridos tanto en los terminales presinápticos de la sinapsis como en su membrana postsináptica, y capaces de persistir desde unos minutos hasta varias semanas. Estos mecanismos resultan tan importantes que merecen una descripción especial.
Memoria a largo plazo.
No existe una delimitación evidente entre los tipos más prolongados de memoria a medio plazo y la auténtica memoria a largo plazo. La distinción es solo de grado. Sin embargo, en general se piensa que la memoria a largo plazo depende de unos cambios estructurales reales sucedidos en las sinapsis, en vez de unos cambios meramente de carácter químico, que potencien o supriman la conducción de las señales. Una vez más, vamos a recordar los experimentos con animales primitivos (donde los sistemas nerviosos son mucho más fáciles de estudiar) que han aportado una inmensa contribución para comprender los posibles mecanismos de la memoria a largo plazo.
El hipocampo favorece el almacenamiento de los recuerdos: la amnesia anterógrada se produce después de lesiones hipocámpicas sostenidas.
El hipocampo es la porción más medial de la corteza en el lóbulo temporal, donde se pliega en un principio siguiendo un sentido medial por debajo del cerebro y después un sentido ascendente hacia la cara interna inferior del ventrículo lateral. Los dos hipocampos se han extirpado para el tratamiento de la epilepsia en unos cuantos pacientes. Este procedimiento no afecta seriamente a la memoria de una persona en lo que atañe a la información almacenada en el cerebro antes de extraer los hipocampos. Sin embargo, una vez realizada la técnica, a partir de entonces prácticamente pierden su capacidad para guardar recuerdos de tipo verbal y simbólico (memoria de tipo declarativo) en la memoria a largo plazo, o incluso en la memoria a medio plazo cuya duración sea superior a unos minutos. Por tanto, estas personas son incapaces de crear nuevos recuerdos a largo plazo con aquellos tipos de información que constituyen los cimientos para la inteligencia. Este trastorno se denomina amnesia anterógrada.